4.1 Ekologie lesa - Úvod

Biosféra, jako živá část planety, místo výskytu živých organismů různého původu i stavby těla a různých ekologických vlastností, zasahuje do většiny dalších zemských "obalů".
Projevy života na naší planetě jsou pozorovány ve všech prostředích mimo souvislý, nenarušený horninový masív. Ode dna oceánů až po nejvyšší vrstvy atmosféry jsou živé organismy nebo materiální doklady jejich existence všudypřítomné. Vzniká tak souvislý objem v rámci planety obsahující život a stopy jeho pobytu, tzv.
biosféra
, oživený prostor planety Země. Pod tímto pojmem se
zároveň shrnuje soubor veškerých živých organizmů v tomto prostoru. Biosféra se v globálním měřítku prolíná s ostatními faktory prostředí, převážně fyzikálního a chemického rázu, tj. s
atmosférou
, vzdušným obalem Země,
hydrosférou
, jež zahrnuje vodu v její plynné, kapalné i tuhé fázi v celoplanetárním měřítku a konečně s
litosférou
, zahrnující horninové prostředí planety. Interakcí těchto fází vzniká další specifické prostředí Země,
pedosféra
, půdní pokryv.
Planeta jeko taková zahrnuje velmi mnohotvárné prostředí s různým typem terénu, různými klimatickými faktory související s tvarem planety a jejím umístění ve vesmíru,
a různým uspořádáním rostlinných a živočišných společenstev. Přesto ale v rámci celé planety existují určité zákonitosti,
obecně platné a ovlivňující vzájemné vztahy organismů mezi sebou, organismů a jejich prostředí a produkční možnosti stanovišť.
Pro funkci globálního geosystému hraje ozvláště důležitou roli vnější energie,
radiační energie Slunce, absorbovaná a transformovaná na povrchu naší planety, do značné míry právě díky živé hmotě.
4.2 Základní pojmy popisující ekosystémy
Živá hmota a její projevy jsou studovány v rámci celé řady specifických vědních oborů. Pro jejich rozlišení je rozhodující metodický přístup a v první řadě úroveň, hladina komplexity, na níž jsou živá hmota a její projevy studovány (Jeník 1995).
Lze tak postupovat od úrovně makromolekul přes struktury membrán, buněčných organel, jednotlivých buněk, pletiv a tkání, orgánů, jednotlivých organismů, jejich populací, společenstev až po úroveň ekosystémů a geosystémů. Na každé z těchto úrovní probíhají interakce mezi živými strukturami a jejich fyzikálním a chemických prostředím, popřípadě i mezi živými strukturami navzájem.
Ekologie
, jako specifická vědní nauka, se pak interakcí živých soustav a jejich prostředí začíná zabývat až od jistých vyšších úrovní, zpravidla od úrovně komplexity organizmů. Lze ji tak definovat jako vědu,
studující vztahy organizmů a jejich skupin k vnějšímu prostředí, čili vztahy mezi živými organizmy (biocenózou) a jejich prostředím(biotopem).
Tomu odpovídá starší pojetí ekologie, kdy převažovalo ryze biologické zaměření studií (Bormann - Likens 1979) na jedince, populace, druhy, skupiny druhů (např. podle stejné funkce v ekosystémech) a společenstva.
V posledním období není předmětem zájmu ekologie pouze živá složky či její části a vztah k okolí. Podle novějšího přístupu
lze ekologii definovat jako vědu studující strukturu, funkci a reprodukci ekosystémů, tj.
nejen vztah jednotlivých individuí a jejich populací, popř. společenstev k jejich prostředí, ale
i materiálové, energetické a informační toky v ekosystémech jako celcích
.
Podle dalších hledisek můžeme rozlišit řadu specializovaných přístupů a disciplín v rámci široce pojaté ekologie:
- autekologii
studující vztahy jednotlivých organismů k jejich konkrétnímu prostředí, - synekologii
zabývající se vztahy skupin organismů a jejich prostředí, - lze studovat ekologii rostlin, živočichů, populací či člověka,
- Fyziologická ekologie rostlin
se zabývá fyziologickými pochody (mj. i primární produkcí, jež je studována detailněji produkční ekologií) v rostlinách ve vztahu k vnějším podmínkám. - Podle umístění zkoumaných ekosystémů můžeme rozlišit terestrickou ekologii, limnologii a oceánologii.
Tropickou ekologii, arktickou ekologii a ekologii aridních území
lze odlišit podle typů fyzicko - geografického prostředí a studované geografické zóny. - Silvologii(nauku o lesním ekosystému), pratologii (nauku o loukách), telmatologii (nauku o rašeliništích)lze konečně odlišit podle typu zkoumaného ekosystému.
Ekosystém
Dalším ústředním pojmem je koncept
ekosystému.
V původním pojetí (Tansley 1935) se jím rozuměl soubor organismů a jejich prostředí v jednotě jakékoli hierarchické úrovně.
V nejobecnější rovině je možno uvažovat každou soustavu, obsahující i jeden živý prvek. Další alternativní pojetí jako ekosystém označují úhrn všech živých organizmů a abiotického prostředí v daném časoprostoru (Jeník 1995),
nebo soubor všech životních forem a jejich projevů probíhajících v uvažovaném období a topograficky vymezeném prostoru.
ON 48 0002 pak popisuje v aktuálně platné definici ekosystém jako:
relativně trvalý dynamický systém souboru organizmů a jejich prostředí (ekotopu) vytvářející určitou stabilitu danou potravními vazbami, resp. koloběhem živin a tokem energie uvnitř této úrovně.
V evropských podmínkách byly definovány alternativní pojmy.
Biogeocenóza
byla popsána V.N. Sukačevem jako část povrchu zemského, na němž biocenóza (fyto-, zoo- a mikrobiocenóza) a jí odpovídající části atmosféry, litosféry a pedosféry i jejich vzájemné vztahy zůstávají stejnorodé, takže tvoří jednotný, vnitřně podmíněný komplex.
Geobiocenóza
definovaná A. Zlatníkem představuje soubor biocenózy (soubor všech organismů daného společenstva) s jejím prostředím.
Jedná se v obou případech o ekosystémový komplex na určité lokalitě, tedy definici ekosystému v užších podmínkách konkrétních stanovišť.
Ekosystémy mohou být uspořádány hierarchicky, tj. jeden může být součástí druhého.
Lze tak postupovat z planetární, globální úrovně na úroveň kontinentální, regionální (na ploše měřitelné zhruba tisíci km2), chorickou (plocha tisíců ha, krajinná úroveň) a topickou (jednotlivé lokality). Je možno definovat subekosystémy, jež jsou součástí ekosystémů vyššího řádu (rákosina v rámci ekosystému rybníka, smrkový porost v rámci lesního komplexu), popřípadě supraekosystémy, zahrnující naopak ekosystémy nižšího řádu (ekosystém krajiny zahrnující lesní porosty).
Ekosystémy mohou být dále uspořádány vedle sebe
, jejich ohraničení bývá přitom z hlediska jejich určení jako systémů otevřených často obtížné. V přírodě mezi ekosystémy existují
hranice konvergentní
(sevřené, ostré) či
divergentní
(pozvolné, difúzní). Ve všech případech je mezi dvěma sousedícími ekosystémy možno vylišit přechodnou zónu čili
ekoton
, zahrnující směs faktorů obou odlišných
typů ekosystémů. Jeho typickým příkladem je lesní plášť. Při plynulé změně půdní, vlhkostní či biotické složky ekosystému, bez vzniku dvou odlišných částí, jedná se o
ekoklinu
Struktura ekosystému
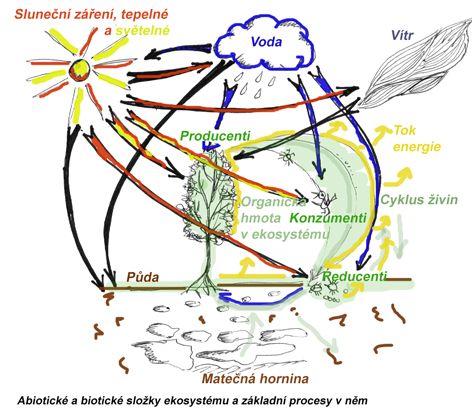
Každý ekosystém je charakterizován souborem abiotických a biotických faktorů jež určují do značné míry jeho možnosti, především z hlediska produkčních možností autotrofních organismů a tím dávají předpoklady pro počátek potravních řetězců a potravních zdrojů pro organismy heterotrofní. Soubor klimatických faktorů dané lokality je označován jako klimatop , režim fyzikálních a chemických činitelů vázaných na substrát jako edafotop (alespoň u terestrických ekosystémů). Klimatop a edafotop tvoří ekotop čili konkrétní stanoviště. Na celkovém režimu jednotlivých stanovišť se velice výrazně podílejí i biotické složky ekosystémů, proto se ve vztahu ke konkrétnímu organismu, populaci či společenstvu hovoří o biotopu, nebo životním prostředí, jež v sobě zahrnuje interakci s abiotickými i biotickými faktory prostředí.
Ekosystémy jsou soustavy dynamické
, tj. proměnlivé v čase.
Změny probíhaly a probíhají v různých časových rozměrech, čili časových horizontech. Lze popsat změny
aktuální
(charakteru události či v intervalu hodin, dnů, roční sezóny a roků),
sekulární
s časomírou staletí a tisíciletí a geohistorické s časovým určením desetitisíců či miliónů let. Sekulární a geohistorické změny jsou předmětem zájmu
paleoekologie
.
Některé změny jsou náhodné, označované jako ekologické fluktuace. Jiné, rytmického charakteru, se opakují s určitou periodou:
cirkadiánní změny
s periodou dne,
lunární
změny korelující s lunárními cykly, či
sezónní
změny opakující se s ročními obdobími.
Biosféra Země se nevratně vyvíjí a spolu s ní procházejí vývojem i ekosystémy různé úrovně a komplexity. Při dostatečně dlouhé době nebo dostatečně intenzivním podnětu dochází ke změnám jejich struktury a funkce evolucí, postupně (z hlediska sekulárních změn), či skokem, náhle.
Základní pojmy popisující vztahy v ekosystému
Ekologická rovnováha označuje stav, kdy se ekosystém udržuje zhruba
konstantní nebo s pravidelným kolísáním.
Je-li dosahován v podmínkách působení rušivých faktorů, je projevem ekologické stability.Obecně se ekosystémy vyznačují tendenci bránit se vnějším vlivům vedoucím k jejich vnitřním změnám, vykazují větší či menší stabilitu.
Stabilita je definována (Míchal 1994) jako schopnost ekologického systému přetrvávat i za působení rušivého vlivu a reprodukovat své podstatné charakteristiky v podmínkách narušování zvenčí.
- minimální změnou za působení rušivého vlivu (rezistence - tzv. odolnost), nebo
- spontánním návratem do výchozího stavu (rezilience - tzv. pružnost)
Ekosystémy se tedy většinou nacházejí ve stavu dynamické rovnováhy.
Přes obměnu složek ekosystémů, růst, vývoj a odumírání organizmů i přes značné energetické, látkové a informační toky ekosystémy zůstává struktura a funkce na zhruba stejné úrovni, nebo prochází v určitých periodách předpovídatelnými a vyrovnanými cykly.
Sukcese
Adaptivní změny vyvolané především činiteli vnějšího (převážně fyzického) prostředí jsou označovány jako exogenní nebo
allogenní sukcese
, změny způsobené vnitřními převážně biotickými procesy (v rámci společenstva) pak jako
endogenní či autogenní sukcese
. V obou případech pak hovoříme o ekologické sukcesi, či jak bylo již uvedeno, o sukcesi.
Relativně stabilní vnější i vnitřní faktory vedou k vytvoření konečného článku sukcesních stadií,
klimaxu
, ve kterém se ustaluje struktura i toky.
V lesnictví a zejména v hospodářských lesích přírodě blízkých je důležitý i pojem
subklimaxu
, kdy je dosažení konečného stadia bráněno hospodářskými (pěstebními) zásahy, přičemž jsou záměrně využívány přírodní procesy vedoucí k formování stabilních lesních porostů.
Popis struktury a funkce ekosystému je záležitost vysoce komplexní a komplikovaná, zatím v úplné podrobnosti neproveditelná. Proto se pracuje velice často s modely ekosystémů, zjednodušeně demonstrujícími hlavní abiotické zdroje a biotické součásti v podobě dílčích bloků (složek, kompartmentů) a dále hlavní přenosy látek, energií a informace v podobě šipek.
Vazby v ekosystému
Stabilizující faktory představují především zpětné vazby v ekosystémech.-
Kladná zpětná vazba
nastává v případě, že jistý proces v ekosystému vede přímo nebo nepřímo k jeho zesílení. Jako příklad lze uvést rozvoj rašeliníku a zamokření lokality. Kladná, pozitivní zpětná vazba je spojena s dynamickým růstem systému, jež se ovšem po dosažení limitů prostředí může zhroutit. -
Negativní zpětná vazba
bývá považována za základ regulace systémů při jejich udržení v rovnovážném stavu - tzv. homeostáze. Jedná se o případ, kdy určitý jev či proces v ekosystému vede při určité intenzitě k podpoře faktorů, potlačujících zpětně daný proces - např. vstup světla do porostu vede k rozvoji přirozeného zmlazení a další růst nárostů vede k zastínění půdy a přerušení fáze obnovy. -
Rovnovážný stav, homeostáza,
je definována (Míchal 1994) "jako soubor principů vedoucích v živých systémech na základě získaných informací ke kompenzaci odchylek vnějšího prostředí, a tím k dynamické rovnováze vnitřního prostředí, které tak nabývá relativní neodvislosti vůči vnějšímu prostředí".
Odum (1977) ji uvádí jako pojem, "kterého se obecně používá pro označení tendence biologických systémů odolávat změnám a setrvávat v rovnovážném stavu".
Na úrovni krajiny uvažuje Míchal (1994) o homeostázi primární, determinované ryze přírodními procesy, a sekundární, určené nahodilou interferencí s lidskou činností.
Působí-li na ekosystém vnější vliv a tento přesahuje obvyklé rozpětí, ekosystém přejde k mobilizaci obranných a nápravných opatření a nachází se ve stavu stresu. Rušivý faktor, stresor, může být přirozeným faktorem v nepřirozených kvantech, nebo faktorem pro ekosystém kvalitativně cizím.
Soubor vnějších podmínek ekosystému se může měnit i způsobem, jež neumožňuje jeho existenci v dosavadní podobě. Rovněž rušivé vlivy, přírodní či antropogenní, mohou způsobit změny nebo zhroucení ekologického systému, jeho disturbanci . Dochází pak k vývoji ekosystému, k sukcesi.
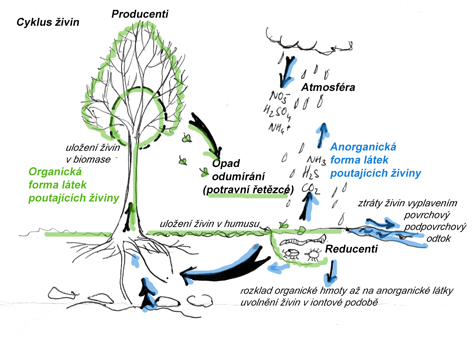
Koloběh minerálních prvků v ekosystému

Koloběh živin v ekosystému spočívá v jejich příjmu rostlinami (v anorganické formě- formě formě iontů rozpuštěných v půdním roztoku), a v uložení v biomase (většinou vazba na organické látky). Přes potravní řetězce a řadu konzumentů se prvky dostávají ve finále do nekromasy (odumřelých zbytků rostlin, živočichů, nebo jejich výlučků) na povrch půdy, kde jsou rozkládány působením mikroorganismů - reducentů - opět až na anorganické látky. V rámci tohoto rozkladu dochází u některých prvků k jejich úniku do atmosféry (dusík, síra) a součástí koloběhu je pak i jejich návrat na zemský povrch- spad z atmosféry. Anorganické látky mohou ve vodě disociovat a ionty daných prvků být přijímány rostlinami, čímž se cyklus uzavírá. Možné ztráty živin z ekosystému spočívají ve vyplavení přes spodní vodu nebo povrchové toky a v možném dálkovém transportu přes atmosféru (jen u prvků , které se do atmosféry dostávají ve formě plynů). Návrat živin do ekosystému je opět ve spadu z atmosféry a při rozkladu matečné horniny. Některé prvky jsou v matečné hornině obsažené ve vyšším množství a velká část jejich objemu je v půdě (např. K), jiné mají těžiště objemu v ekosystému v atmosféře nebo biomase (např N, P). Aby mohl ekosystém fungovat dlouhodobě, musí jít o uzavřený cyklus, ve kterém jsou ztráty dostatečně nahrazovány a nedochází k dlouhodobému úytku žádné ze základních živin.
Koloběh živin těsně souvisí s tokem energie - v počáteční fázi poutání sluneční energie autotrofními organismy (producenty) v procesu fotosyntézy je stavba biomasy těsně spjata s dostupností minerálních živin pro tuto stavbu nutných. V průběhu života organismů je však tato energie, poutaná v organických látkách, neustále využívána (prodýchávání, tepelné ztráty, využití na tvorbu vlastní biomasy) a při rozkladu odumřelých těl až na anorganické látky jsou uvolněny její poslední zbytky. Tok energie tudíž není na rozdíl od koloběhu živin cyklem, ale jednosměrným tokem, ve kterém je nutný neustálý přísun této energie ze slunce.
Obsah prvků v rostlinách
Hlavní biogenní prvky, tvořící většinu sušiny- ve formě organických látek, jsou C - 45%, O- 45%, H - 6%
Látky, které jsou rovněž obsažené v sušině rostlin (organismů) ale při spalování (za nižších teplot)) se neuvolňují ve formě oxidů
se nazývají popeloviny (starý název).
- listnaté stromy : listy - 3-4% , dřevo - 0,5%, kůra - 3-8%
- Jehličnaté stromy: listy - 4%, dřevo 0,4 % , kůra - 3-4%
Ztráty živin z biomasy :
Minerální prvky, které rostlina již přijala přes kořeny a v rámci transpiračního proudu odvedla do asimilačního aparátu,
ve valné většině využívá. Avšak může docházet i k jejich ztrátám.Nejpodstatnější (i když relativně nepravidelné) může být za jistých okolností- zejména u jehličnanů nebo neopadavých listnáčů-
defoliace hmyzem (u jehličnatých může ztráta převýšit roční čerpání živin z půdy. Běžná je ztráta živin opadem listů, ve kterých dochází k poutání živin na několik let.
možné je i vyluhování dešťovou vodou z listů, kůry, kořenů (Na zřídka,- K, Ca, Mg, Mn ve větším množství,řádově v kg/ha).
Fenolické látky vyluhované dešťovou vodou napomáhají podzolizaci.
Makroelementy
Prvky, které se v organismu vyskytují řádově v procentech, případně desetině, minimálně setině procenta. Tvoří významnou stavební součást látek s různou funkcí v organismu - stavebních (bílkoviny, DNA), regulačních (enzymy, fytohormony), strukturních (fosfolipidy), energetických (ATP), pomáhají při udržení homeostázy vnitřního prostředí (regulace sycení buněk vodou)
Dusík
Obsah v rostlině:
0,5-5% sušiny , prům. 1,5 % (byliny 2-4%, listy stromů 1-4%, jehlice 1-2%)
Význam v rostlině: základní složka protoplazmy, bílkovin a enzymů
Hromadí se: listy, mladé prýty, pupeny, semena, zásobní orgány.
Příznaky nedostatku: omezení tvorby stavebních a funkčních
bílkovin - trpasličí vzrůst - omezený hmotnostní přírůstek,
větší podíl kořenů, neplodnost květů, předčasné žloutnutí listů, listy slabší,
tenčí, světlejší, menší buňky se ztloustlými stěnami, omezení tvorby listů
vede k omezení fotosyntézy a tvorby nadzemní biomasy
většinou to vede k rychlejšímu dozrávání a kratší vegetační době, ale zároveň
ke snížení kvality semen.
Nadbytek - nevyzrálá pletiva, která mohou namrzat
Příjem ve formě: NH4+ z humusu a koloidů,
NO3- z roztoku,
Koloběh v půdě: Ve formě odumřelé biomasy je vázán v různých typech organických látek, ze kterých je většinou rozkladem uvolňován jako amonný kationt.
Mikroorganismy - návrat organicky vázaného dusíku do výchozích anorg. sloučenin.
V půdě dochází k dalším transformacím forem dusíku: z původního amonného iontu uvolněného z nekromasy oxidací nitritačními bakteriemi na dusitany a
nitratačními bakteriemi na dusičnany v procesu nitrifikace (nutné vyšší teploty a vlhkost půdy, ne příliš kyselé pH).
V půdě existují i denitrifikační bakterie, které naopak formy dusíku redukují, což vede k úniku oxidů dusíku z půdy do atmosféry.
Dusík v atmosféře v plynné formě N2 je pro rostliny nepřijatelný, ale specializované typy organismů - bakterií- ho dokáží poutat i v této nformě.
Nesymbiotické fixace dusíku - se účastní volně žijící půdní bakterie Clostridium pasteurianum - anaerob.,
Azotobacter chroococcum - aerobní, sinice - Nostoc, Anabaena, Calothrix,
Redukčním štěpením N2 je (katalyzováno nitrogenázovým systémem
- Mo, Fe- nutný dostatek těchto prvků) energie dodávána respirací, (opt. teplota 20-30 °C, min. 0 °C ,
dostatek vlhkosti a rozloženého humusu)
Produkce - arktida 0,1-2 kg N/ha/rok, rýžoviště 50-70 kg N/ha/rok, lesní půdy 1-50 kg N/ha/rok
Symbiotická fixace spočívá v symbiotickém vztahu bakterie a rostliny, přičemž potravou pro bakterie jsou glycidy z buněk rostliny
a výhoda pro rostlinu spočívá v lepší přístupnosti dusíku.
Rhizobium - Fabales, Azotobacter - C4 trávy,
Frankia - Alnus, Myrica, Hippophae, Eleagnus, Casuarina, sinice - mechy, lišejníky, kapraďorosty..)
Fosfor
Obsah v rostlině: 0,1-1% , prům. 0,2% sušiny
Význam v rostlině:základní metabolismus a syntézy, fosforylace - fotosyntéza, DNA
Hromadí se: více v reprodukčních, než ve vegetativních orgánech, v květech až 1,5% v sušině, v pylových zrnech 2-3%, semena 0,5-1%
Příznaky nedostatku:většinou latentně - nízký obsah v rostlině - snížená úroveň biochem. procesů, především se přestávají tvořit cukry a bílkoviny
výrazněji se projeví za chladného, nebo suchého počasí. Dále poruchy reprodukce ( zpožděné kvetení, snížené zakládání květů, špatná tvorba semen),
zakrslost, tmavě zelené, nebo bronzově fialové zbarvení, zasýchání špiček jehlic u jehličnanů, omezený růst letorostů.
Dobrá výživa - urychluje vyzrávání dřeva, zkracuje vegetační období, snižuje negativní účinky přehnojení dusíkemNadbytek - pouze krátkodobě ( vyplaví se) - může způsobit druhotný nedostatek zinku
Příjem ve formě: H2PO42- a
HPO4- - hůře rozpustný zejm. ve vápenatých půdách.
Koloběh v půdě: uvolňuje se z nerostu apatitu, v půdě je ve formě málo rozpustných fosforečnanů
a lépe rozpustných hydrogenfosforečnanů Ca, Mg, Al, Fe.
Z organické hmoty se uvolňuje hydrolýzou.
Ve velmi kyselých půdách se sráží ve spodině profilu.
Draslík
Obsah v rostlině:0,2 - 5% , prům. 1% sušiny
Význam v rostlině:podporuje hydrataci pletiv a osmoregulaci, aktivace enzymů
ve fotosyntéze, antagonismus s Ca, synergismus s Na a NH4
Hromadí se : meristémy (kambium), mladá pletiva, intenzivní metabolismus, z
listů může být vymýván srážkami.
Příznaky nedostatku:
zasýchání vrcholků, poruchy vodního metabolismu, předčasný
opad starých ročníků jehlic, zvlněné a zasychající okraje starých listů,
kořenová hniloba, zakrnělé rostliny, malá semena případně zásobní hlízy. Dostatečná výživa - dobré vyzrávání a mrazuvzdornost pletiv, cukernatost plodů
Nadbytek - poruchy ve výživě vápníkem, sytě zelené listy, bujný růst, odumírající starší listy
Příjem ve formě:K+, pro příjem jsou nepříznivé podmínky v odbdobí sucha a chladu.
Koloběh v půdě: složkou hornin a minerálů (živce), uvolňován zvětráváním,
váže se na půdní koloidy a je rozpuštěn v půdním roztoku (je snadno pohyblivý),
optimální obsah je 3-4% z celkové sorpční apacity, při vyšším obsahu je
vyplavován do půdního roztoku v nadbytku v půdním roztoku brání příjmu Na, Mg, Ca
Vápník
Obsah v rostlině:0,1 - 5%, prům. 0,5% sušiny
Význam v rostlině:regulace hydratace (antagonisté K, Mg), aktivace amylázy,
ATPázy, regulace dlouživého růstu
Hromadí se : listy, kůra, starší pletiva, (buněčná stěna - střední lamela).
Příznaky nedostatku:
poruchy při dělivém růstu ( malé buňky), zasýchání vrcholků a
listů (místa nejintenzivnějšího prodlužovacího růstu- nevytvoří se dostatečně
kvalitní buněčné stěny), zbržděný růst kořenů.Dostatek - napomáhá vyzrávání dřeva
Příjem ve formě:Ca2+, deficitní ve velmi kyselých půdách, v rostlině málo
pohyblivý, příjem K+ve vlhku a teplu působí jako antagonista příjmu Ca
Koloběh v půdě: uvolňován z hornin ( vápence, dolomity). v rozpustné formě
jako hydrogenuhličitan, za přítomnosti CO2 se sráží na nerozpustný uhličitan,
v sorpčním komplexu by měl tvořit 60-80%
Hořčík
Obsah v rostlině:0,1-1% , prům. 0,3% sušiny
Význam v rostlině:součást chlorofylu, bílkoviny buněčného jádra, střední lamela
buněčné stěny, regulace hydratace (antagonismus Ca), fotosyntéza - přenos fosfátů, (synergisté Mn, Zn)
Hromadí se : Listy, Příznaky nedostatku:
zakrslý růst, chloróza mezi žilnatinou starých listů, nepravidelně
zprohýbané listy, při případném odumírání, purpurové zbarvení listů jako při nedostatku P
Příjem ve formě:Mg2+, deficitní v kyselých půdách, kdy je deficit doprovázen nadbytkem Mn
Síra
Obsah v rostlině:0,1 - 1%, prům. 0,1% sušiny
Význam v rostlině: složka protoplazmy, aminokyselin, enzymů
Hromadí se : listy, semena, Příznaky nedostatku: mezižeberní chloróza mladých listů, nedostatek Mg ve starších listech
Příjem ve formě:
SO42-
Koloběh v půdě: Síra je uvolňována z hornin a nerostů (pyrit, chalkopyrit) ve formě síranů (okyselování půdy).
V organismech v AK (cystein), mineralizací uvolňován sirovodík, ten oxiduje za vzniku síranů.
Jako prvek, který se snadno oxiduje a tudíž je využitelný pro přenos chemické energie, je využíván i specializovanými typy organismů:
- příkladem jsou sirné bakterie, které síru využívají při životních procesech místo kyslíku.
Železo
Obsah v rostlině:0,02 - 0,001% sušiny
Význam v rostlině: redoxní reakce, základní metabolismus, metabolismus dusíku,
syntéza chlorofylu, (obsažen v cytochromu, ferredoxinu)
Hromadí se : listy,
Příznaky nedostatku: slámově žluté mezižeberní chlorózy, v extrému bílé zbarvení
mladých listů, potlačená tvorba apikálních pupenů
Příjem ve formě: chelátů, v redukovaném stavu Fe2+, transport v rostlině špatný, ve vápenatých zásaditých půdách pevně vázáno
Koloběh v půdě:
výskyt v oxidech, hydroxidech, fosforečnanech, jílových minerálech.
Uvolňuje se zvětráváním redoxní reakcí půdních mikroorganismů (Thiobacillus ferrooxidans),
kteří využívají uvolněnou energii pro život.
H +vznikající produkcí anaerobních bakterií (Clostridium, Pseudomonas) redukuje Fe3+
(zrzavočervené zbarvení) na Fe2+ ----zelenavé zbarvení půd
V alkalickém půdním roztoku snadno tvoří nerozpustné sraženiny (půdy s přebytkem vápníku).
Mikroelementy
Prvky, jejichž obsah v rostlinném těle je tak nízký, že se udává v jednotkách ppm (mg/kg)
Prvky (často kovové) které jsou často součástí enzymů, barviv, účastní se redoxních reakcí a přenosů energie.
V rostlině se vyskytují ve velmi malém množství ale mohou mít relativně
velký vliv na intenzitu a průběh metabolických reakcí, typická je funkce regulátorů aktivity enzymů.
Charakteristická je špatná pohyblivost v rostlině (kromě Cl).
Měď
Obsah v rostlině:6 ppm
Význam v rostlině:základní metabolismus (fotosyntéza, oxidázy - fenoloxidáza,barviva
- plastocyanin) metabolismus dusíku, druhotný metabolismus,
Hromadí se : zdřevnatělé nadzemní stonky,
Příznaky nedostatku: zasýchání vrcholků, zkroucené listy, chlorotické skvrny na mladých listech
Příjem ve formě: Cu2+, Cu - chelátů, silná vazba na humus,
v kyselém prostředí pohyblivější
Zinek
Obsah v rostlině:20 ppm
Význam v rostlině:
tvorba chlorofylu, aktivace enzymů, základní metabolismus
- dehydrogenázy, rozklad bílkovin, biosyntéza růstových regulátorů
Hromadí se :kořeny, prýty, reprodukční orgány - v rostlině pomalý pohyb
Příznaky nedostatku:zakrslý vzrůst, bílo-zelené zbarvení starších listů, poruchy plodnosti
Příjem ve formě:Zn2+, Zn - chelátů, v kyselém prostředí dostupnější
Molybden
Obsah v rostlině:0,1 ppm
Význam v rostlině:fixace dusíku, metabolismus fosforu, absorpce a translokace železa,
metaloorganické sloučeniny a enzymy
Příznaky nedostatku:poruchy růstu, deformace prýtu, hnědnutí listových okrajů, chlorotické drobné skvrny
Příjem ve formě:
MoO42+, v bazickém prostředí dostupnější
Mangan
Obsah v rostlině:50 ppm
Význam v rostlině: základní metabolismus ( oxidázy, fotosyntéza,
přenos fosfátů, metaloorganické sloučeniny), stabilizace struktury
chloroplastů, metabolismus dusíku, nukleových kyselin, synergisté : Mg, Zn
Příznaky nedostatku: inhibice růstu, chlorózy a nekrózy mladých listů, opad listů
Příjem ve formě: Mn2+, Mn - chelát, v kyselém prostředí dostupnější,
pro rostliny přístupný jen Mn2+, Mn4+ve vodě nerozpustná forma.
Bór
Obsah v rostlině:20 ppm
Význam v rostlině: metabolismus a transport glycidů, metabolismus fenolů,
ativace růstových regulátorů (růst pylových láček)
Hromadí se :listy, vrcholky prýtů a příznaky nedostatku jsou poruchy růstu, nekrózy meristémů, poruchy plodnosti,
málo se větvící kořeny, nadměrná tvorba korku, skvrnitost plodů u jabloní
Příjem ve formě: HBO32+ ,
H2BO3+v kyselém prostředí je dostupnější
Chlór
Obsah v rostlině:100 ppm
Význam v rostlině:zvyšování hydratace pletiv, aktivace enzymů ve fotosyntéze
Hromadí sev listech, příznaky nedostatku jsou tedy zkroucené listy, ztloustlé kořeny.
Příjem ve formě: Cl-
Stopové prvky
Sodík, Selen, .....Kobalt (symbiotická fixace dusíku)4.3 Jednotlivé složky ekosystému
-
Z hlediska strukturního
lze vylišit složku nadzemní části vegetace, podzemní části vegetace, v rámci které můžeme rozlišovat složku dřevinnou (stromovou, keřovou) a bylinnou. Dále se vylišuje složka nadložního humusu, vzdušného prostředí a půdy, eventuálně i složka živočišná. Přístupů je několik a výsledkem je např. vylišení jednotlivých složek, etáží a vrstev (v korunové části lesního ekosystému). -
Z hlediska funkčního
rozlišujeme kromě abioticky podmíněných vstupů a výstupů (srážky, spady, půdní odtok, eroze) i část biotickou a v ní několik skupin organizmů z hlediska jejich podílu na energetických a látkových vazbách v ekosystémech (producenty, konzumenty, destruenty, reducenty).
4.4 Biotická část ekosystému

Zahrnuje veškeré živé organismy jako součást ekosystému. Je to součást velmi dynamická, s rychlým kolísáním a změnami (ve srovnání se složkou abiotickou) a výrazným tokem a transformacemi energie a využitím a koloběhem živin.
Potravní řetězce
Organismy v reálných podmínkách vytvářejí potravní řetězce nebo při komplikovanějších systémech častěji potravní sítě, popisující tok vytvořené organické hmoty a v ní vázané energie a současně koloběh živin v ekosystému
Rozdělení organismů z hlediska funkčního zařazení v ekosystému
- producenti, autotrofní organismy
podílející se na primární produkci
(zelené rostliny, cyanobaktérie, chemautotrofní organismy) - konzumenti, heterotrofní organismy
(živočichové, houby, protista, saprofytické a parazitické rostliny, prokaryota), mezi kterými je možno dále vylišovat např. herbivory, živící se rostlinnou potravou (jinak konzumenti 1. řádu), predátory, živící se živočišnou potravou (konzumenti 2. a vyšších řádů), parazity a saprofágní-detritofágní-nekrofágní destruenty, živící se odumřelou organickou hmotou. - destruenti (dekompozitoři)
někdy bývají zařazováni mezi konzumenty a to konzumenty saprofágní- živí se odumřelými částmi rostlin a živočichů a řadíme sem různé druhy hmyzu. - reducenti (rozkladači)
baktérie a houby rozkládající organické látky až na minerální součásti
Blíže se rozlišují dva typy energo-materiálových řetězců (sítí):
- tzv. krátký detritofágní řetězec (síť),
zahrnující producenty, destruenty a reducenty,
který je součástí všech fungujících ekosystémů. V lesních ekosystémech je přitom dominantní, většina primární produkce sleduje posloupnost opad-fragmentace-postupný rozklad-mineralizace. - tzv. dlouhý pastevně kořistnický řetězec
(síť),
zahrnující producenty, síť konzumentů, destruenty a reducenty
Tyto řetězce se týkají především biomasy (do které patří i mrtvé části živých organismů - dřevo, chlupy, drápy...), ale i nekromasy, ze které jsou pomocí reducentů uvolňovány živiny, které se tak mohou vracet zpět do koloběhu. Patřísem i pojem gilda - skupina organismů s různým původem ale společným způsobem obživy (př. králík a ovce).
vzájemné vztahy mezi druhy
koevoluce
- proces, při němž členové dvou druhů recipročně podporují síly přirozeného výběru, kterými na sebe navzájem působí (paraziti a hostitelé např.)
amensalismus
- jeden druh působí nepříznivě na druhý, v opačném směru vztah neutrální
koexistence
- soužití dvou navzájem se nevylučujících druhů na stejném stanovišti asymetrická konkurence - konkurence dvou druhů, při níž je jeden postihován více než druhý
mutualismus
- (vzájemnost) - vztah, kdy interakce mezi jedinci dvou/více druhů pozitivně ovlivňuje jejich růst nebo velikost populací. (př. mykorhiza - pevné mutualistické spojení mezi kořeny rostliny a houbou)
fakultativní mutualismus
- vztah, kdy jeden nebo oba druhy mohou přežívat i při absenci druhého druhu, ale pokud jsou spolu, tak se vzájemně pozitivně ovlivňují.
komensalismus
- interakce, kdy jeden organismus působí kladně na jiný a ten naopak nijak nepůsobí na organismus první.
vzájemný antagonismus
- negativní reciproční ovlivňování dvou druhů (mezidruhová konkurence, nebo vzájemná predace)
zdánlivá konkurence
- interakce, kdy na sebe druhy navzájem negativně působí tím, že pozitivně ovlivňují společného predátora
konkurence
- vztah dvou (nebo více ) druhů, kdy činnost jednoho organismu nebo druhu potlačuje rychlost růstu nebo natalitu druhého druhu, nebo zvyšuje jeho mortalitu. (Možnost vytlačení jednoho druhu ze stanoviště jiným druhem). konkurence možná přerůstáním (brání v zásobením světlem, živinami).
endobiont
- organismus žijící uvnitř buněk hostitele, nezpůsobující zjevnou škodu
hemiparazit
- rostlina schopná fotosyntézy, uzpůsobená čerpat vodu a /nebo/ minerální živiny z kmene nebo kořenů hostitele
holoparazit
- úplný parazit - parazitické rostliny bez chlorofylu, výživa plně závislá na hostiteli
nekroparazit
- parazit, který usmrcuje svého hostitele a dále žije z jeho mrtvé hmoty
parazit
- organismus získávající své živiny z hostitele, kterého sice poškozuje, ale nepůsobí mu okamžitou smrt
spojené výkyvy (oscilace)
- závislé výkyvy početnosti dvou druhů, kdy nízká početnost kořisti vede k nízké početnosti predátora a to dále vyvolává vysokou početnost kořisti a následně i predátora atd.
vzájemná interference
- vzájemné působení predátorů, které roste s hustotou jejich výskytu a ztěžuje jim příjem potravy
Životní formy rostlin (podle Raunkiaera)
je lze rozdělit na:Fanerofyty
- přetrvávající pupeny či vegetační vrcholy na výhoncích rozprostřených v nadzemním prostoru (vždyzelené a opadavé dřeviny)
Chamaefyty
- přetrvávající pupeny, nebo vegetační vrcholy na výhoncích nízko u země (rostliny plazivé, nebo s polštářovitým růstem, nebo keříčkovité, jejichž výhonky od vrcholku odumírají až k části nesoucí přežívající pupeny)
Hemikryptofyty
- přetrvávající pupeny, nebo vegetační vrcholy jsou na povrchu půdy (růžicovitě rostoucí rostliny)
Kryptofyty
- pupeny, nebo vegetační vrcholy jsou skryté v půdě (nebo vodě) - (rostliny s oddenky, cibulemi, hlízami, vodní nebo mokřadní rostliny)
Terofyty
- rostliny s krátkým životním cyklem během jedné vegetační sezóny, od semene k semeni (také označovány jako efemerní rostliny)
Halofyty
- rostliny schopné žít ve velmi slané vodě, nebo půdě
Populace a společenstva
Jako populace jsou přitom označovány biotické systémy vytvářené vzájemně podobnými jedinci téhož druhu, kteří si ve společném prostoru a ve společném čase při pohlavním rozmnožování předávají společné znaky.
Biocenóza, cenóza či společenstvo je pak biotický systém, který zahrnuje synergicky propojené
populace více druhů v rámci určitého časoprostoru.
Podle životních forem, specializace odborníků a systematických skupin lze pak hovořit o fytocenózách, bryocenózách, mykocenózách a zoocenozách. Strukturu společenstev je možno členit i s ohledem na podobnou životní strategii - synusie, na stáří - kohorty, popřípadě na nároky na potravní zdroje - guildy.
Pojmy charakterizující společenstva a ekosystémy
Evolučně stabilní strategie- strategie kterou přijme většina populace, nemůže být vylepšena jinou strategií a patrně jí potvrdí přirozený výběr. Podstatou strategie, je přizpůsobení ke dvěma možným typům nepříznivých vlivů, a to je dlouhodobý stres (suboptimální podmínky), nebo disturbance (narušení), které jsou naopak silné, nepředvídatelné a krátkodobé. Z hlediska snahy vyrovnat se s těmito možnými vlivy rozlišujeme
- K - strategie (konkurenční)
- na stanovištích z hlediska času trvalých, nebo předvídatelně sezónních, vznikne silná populace, početně na úrovni blízké úživnosti prostředí. Existují zde silné (i vnitrodruhové) konkurenční vztahy, dlouhověkost, pozdní plodnost, výraznější přizpůsobení konkrétnímu biotopu, vyšší důraz na konkurenceschopnost a kvalitu (méně početného) potomstva. Tato strategie umožňuje také určité tlumení menších výkyvů v prostředí, díky hromadění energie v zásobních orgánech rostlin (velká semena, biomasa...). - R - strategie (ruderální)
- na stanovištích s nepředvídatelně, nebo rychle vznikajícími silnými disturbancemi. Podstatou strategie je soustředení na velké množství i méně kvalitního potomstva, rychlé množení a šíření populace, relativně malou velikost populace. Prostředkem je soustředení energie na množení, součástí čehož je časná reprodukce, krátkověkost, menší tvorba biomasy, schopnost přizpůsobit se změně prostředí a nízké nároky na podmínky prostředí. Typickými R stratégy jsou pionýrské dřeviny, plevele... - S - strategie (stres snášející)
- na stanovištích s inztenzivní mírou nějakého typu stresu (klimatické nebo půdní extrémy- extrémní sucho, nedostatek živin, zasolení...). Typická pro ně je nízká tvorba biomasy, pomalý růst, relativně nižší plodnost a velmi silné adaptace na konkrétní prostředí (v prostředí nízké konkurence). Nepravidelné a silné narušení stanovišť s stratégové snášejí špatně.
Běžná strategie na slaništích, rašeliništích, vřesovištích...
Společenstvo
- Druhy, které se vyskytují společně v prostoru a čase (často jsou mezi nimi vzájemné vztahy a společně jsou vázány na určitý typ prostředí)
Dynamicky labilní
- společenstvo stabilní jen v rámci relativně úzkého spektra podmínek prostředí
Dynamicky stabilní
- stabilní v rámci širokého spektra podmínek prostředí
Nestabilní rovnováha
- stav populací, nebo zdrojů, kdy malý výkyv vede k výkyvu rozsáhlejšímu
Nosná kapacita prostředí
- maximální velikost populace, která může v daném prostředí neomezeně dlouho existovat: vnitrodruhová konkurence při ní redukuje čistou růstovou rychlost populace na nulu.
Závislost na hustotě - při rostoucí hustotě má populace tendenci zvyšovat mortalitu a snižovat natalitu a rychlost růstu.
Teorie jednoho klimaxu
- Koncepce předpokládající že všechny sukcesní řady v
dané oblasti vedou k jednomu charakteristickému konečnému společenstvu
Teorie více vrcholů
- Koncepce, podle které sukcese vede k jednomu z více možných cílových společenstev podle podmínek prostředí
4.5 Abiotická část ekosystému
Zahrnuje neživou část ekosystému - a to složky, které umožňují existenci živých organismů tím, že formují jejich prostředí, určují dostupnost vody, živin - chemických prvků nezbytných pro život organismů, dostupnost energie charakterizující okolní prostředí ale i potravní zdroje.
4.6 Sluneční záření

Sluneční záření má dvě základní formy, které ovlivňují výrazněji charakter a fungování ekosystémů a které z těchto důvodů odlišujeme. Jedná se o záření světelné a tepelné. Sluneční záření je abiotickým faktorem, který má zásadní vliv na biotickou složku ekosystémů - jeho energie, poutaná producenty, je hlavním zdrojem pro vytvoření organických látek - a tím zprostředkovaně potravním zdrojem všech živých organismů v ekosystému. Světlo je současně faktorem, podle kterého se orientuje značná část živočichů, ale i rostlin, které podle něj jsou schopné určit směr růstu, ale i část roku, nebo vegetačního období. Teplo rovněž ovlivňuje produkční možnosti rostlin a možnosti přežití a aktivity živočichů a to i půdních. Rozklad organické hmoty v půdě, jako součást koloběhu živin, je určován především podmínkami pro možnosti života mikroorganismů. Sluneční záření ovlivňuje i abiotické složky ekosystému: vodní režim, proudění vzduchu, dostupnost živin v půdě- zde však hraje hlavní roli záření tepelné, určující optimální podmínky pro chemické procesy v půdě a jeho extrémy ovlivňující zvětrávání matečné horniny. Tepelné záření ovlivňuje výpar vody do atmosféry a formování oblaků, které naopak snižují intenzitu záření na zemském povrchu. Proudění větru je dáno pohybem vzdušných mas na základě rozdílných teplot.
Z hlediska hodnocení tohoto faktoru v ekosystému jsou sice důležité průměrné roční teploty, délka slunečního svitu, ale především kolísání těchto hodnot, jejich výkyvy a rozdíly mezi dnem a nocí a mezi jednotlivými částmi roku, nebo vegetační doby. Limitujícím faktorem je sluneční záření- tepelné- z hlediska teplotního minima nebo maxima a sumy teplot v průběhu vegetační doby, z hlediska světelného záření pak dostupnost světla především v konkurenci rostlinných druhů. Teplota je limitujícím faktorem v biomech s teplotami příliš vysokými a nízkou dostupností vody (pouště, polopouště) a v biomech s tak nízkými teplotami v určité části roku, že to omezuje růst rostlin a určuje délku vegetační doby (lesy mírného pásu, chladné stepi, tajga, tundra).
Sluneční, nebo solární konstanta 1354 W.m-2, představuje intenzitu záření nad hranicí atmosféry,
jež je průchodem ovzduším snížena pohlcováním, absorpcí a rozptylem, difúzí.
Z hlediska konkrétní plošky na zemském povrchu a její energetické bilance pak můžeme hovořit o
následujících bilančních složkách:
globálním záření jako sumě zářivé energie dopadající na plochu,
skládající se z přímého záření (insolace)
a rozptýleného (difúzního) záření,
a doplněného o odražené záření (albedo)
a tepelného vyzařování místa i atmosféry.
Záření podléhá sezónním změnám v rámci rotace Země (den-noc, diurnální), oběhu Země kolem Slunce
(roční, sezónní, cirkadiánní periodicita) a proměnlivé sluneční aktivity.
Také poloha na zemském povrchu (zeměpisná šířka) určuje rozhodujícím způsobem dynamiku a množství záření.
Ekologický význam záření
a jeho jednotlivých složek může být dosti rozdílný.
Difúzní záření může být rostlinami využito v celém rozsahu,
přímé a příliš intenzivní záření jim může i škodit.
Záření kratších vlnových délek,
i záření ultrafialové, má letální účinky z hlediska poškození citlivějších tkání živých organismů i genetické informace buněk. Nicméně je téměř zcela pohlcováno atmosférou,
v závislosti na její mocnosti a množství a typu v ní obsažených částic.
Na zemský povrch ho proniká jen malá část, i ta však některé formy života značně ohrožuje.
Rostliny se proti nepříznivým vlivům všech forem záření brání
(ochlupení, vosková vrstva).
Infračervené záření představuje tepelné záření objektů, včetně atmosféry.
Je důležité pro udržení pro život příznivého rozmezí teplot.
Celkově jde o složené záření skládající se z fotonů pohybujících se na různých vlnových délkách v rámci celé škály vyzařované Sluncem, tj. v rámci celého slunečního spektra. Většina energie záření časem degraduje na záření tepelné, část energie se mění na jiné formy (energie poutaná chemicky (fotosyntéza), fyzikálně (výparem)apod..
Rozdělení spektra slunečního záření
- Ultrafialové záření (UV)
vlnová délka 290 - 380 nm
podíl z celkového záření (0-4 %) závisí na vzdálenosti od zemského povrchu, směrem k vrchním vrstvám atmosféry jeho intenzita stoupá
fotosyntetický i tepelný účinek je nevýznamný, naopak má významné účinky fotodestrukční - Viditelné záření
vlnová délka 380 - 710 nm
podíl z celkového záření (21-46 %)
krom velmi významného účinku fotosyntetického má významný účinek i v oblasti tepelné - Infračervené záření - IR
vlnová délka 710 - 4 000 nm
podíl z celkového záření (50 - 79 %)
významné pedevším v tepelné oblasti, kdy vyzařování tepla hraje důležitou roli v energetické bilanci živočichů, zejména teplokrevných. Jeho suma je dána přímým vstupem do atmosféry se slunečním zářením a tepelným vyzařováním objektů, jež přijaly energii jiných vlnových délek. - Dlouhovlnné záření
vlnová délka 4 000 - 100 000 nm
podíl z celkového záření (50 - 79 %)
významné pedevším v tepelné oblasti
Fotosynteticky aktivní - světelné- záření
(je označováno jako FAR)
- v oboru viditelného světla, 450 - 760 nm
má i zásadní vliv pro fotosyntézu, je využíváno primárními producenty a převáděno na chemickou
energii organických vazeb. Je zásadním vstupem pro vytváření produkce lesního ekosystému,
jakož i naprosté většiny dalších ekosystémů.
Les a světelné záření

Lesní ekosystém, hlavně korunová vrstva, má zásadní vliv na modifikaci procházejícího záření
a na radiační bilanci lokality.
Část zářivé energie je odražena zpět do ovzduší (albedo, reflexe)
Další část je pohlcena biomasou
(absorpce)
a část proniká listy a korunovou vrstvou (transmise).
Reflexe a transmise představují
záření využitelné pro další fotosynteticky aktivní orgány a organizmy. Množství
světla pronikajícího do nitra porostu (porostního prostředí) je oproti množství světla nad korunami stromů
značně sníženo a je změněna jeho vlnová délka (chybí především modrá a červená část spektra).
Pokles ozářenosti v rámci korunového prostoru klesá nelineárně, na hodnotu vylučující existenci asimilačního aparátu rostlin - kompenzační bod fotosyntézy (zhruba ve spodní třetině koruny), potom se až k půdnímu povrchu příliš nemění. Na ozářenost porostního prostředí má vliv především druh dřeviny (inztenzita světla na povrchu půdy v porostech buku a smrku je zhruba 3-7%, u borovice a břízy 10-20%) a hustota porostu. Také pěstební zásahy ovlivňující počet stromů na stanovišti mění výrazně poměry v porostech lesních dřevin, což má vliv na celý ekosystém lesa. Nicméně efekt těchto zásahů, pokud nemají vysokou intenzitu, je většinou jen krátkodobý a poměrně rychle odezní, neboť během krátkého časového intervalu lesní dřeviny uvolněný prostor rychle zaplní.
Množství světla v porostech lesních dřevin (Klimo et al. 1994) ve srovnání s % světla na volné ploše se podle
ročního období (zejména u opadavých listnáčů) může měnit až o několik desítek procent,
u stálezelených je tento efekt podstatně menší. Řádově se intenzita světla mimo vegetační období
u opadavých listnáčů pohybuje okolo 60-80% intenzity mimo porost.
Rozdíly mezi porosty různých dřevin se spíše týkají velikosti listové plochy
a typu asimilačního aparátu, než toho, zda se jedná o dřeviny listnaté, nebo jehličnaté.
Tepelné záření

Teplota je určována do značné míry právě zářením, tj. radiačním režimem a dále tepelnými vlastnostmi prostředí (pohlcováním, akumulací, vedením a vyzařováním), a sezónní a periodické změny teploty lokality jsou pak označovány jako teplotní režim.
Jednotlivé organismy, druhy a celá společenstva jsou vázány na určité rozpětí teplot, v němž mohou vegetovat a reprodukovat se, lze nalézt teplotu z určitého hlediska pro jednotlivé projevy optimální a stejně tak lze určit i extrémy, teploty limitní, stejně významné jako teploty pro daný projev života průměrné. Teploty nutné pro zachování života rostliny se pohybují od -5 do +55 °C.
Teplota vzduchu se měří ve výšce 2 m nad zemským povrchem (ve stínu, ve standardních podmíkách, v meteorologické budce). Zpravidla se měří: denní minimální teplota,denní maximální teplota a průměrná denní teplota)
Průměrná denní teplota
je průměr hodnot teploty vzduchu naměřených v: 7:00, 14:00 a 21:00 hod.Výpočet : t(průměrná denní) = (t(7:00) + t(14:00) + 2t(21:00))/4
Délka vegetační doby
vychází z průměrné denní teploty a počítá se jako dlouhodobý průměrný počet dnů v roce, kdy je průměrná denní teplota na daném stanovišti vyšší, než 10oC.Pro asimilaci je u vyšších rostlin jako
- tepelné minimum uvažována hranice 1 - 5 °C
- optimum je zpravidla mezi 20 - 30 °C
- horní limit pak představují teploty mezi 30 - 50 °C
Růstově a produkčně jsou rostliny aktivní při teplotách mezi +5 až +40 °C.
Existují rozdíly mezi teplotou vzduchu a teplotou rostliny samotné.
Při přímém oslunění může být teplota povrchu rostlinného těla o 2-8 °C
vyšší než teplota vzduchu.
Citlivá jsou zejména mladá vývojová stádia dřevin (některé typy padání semenáčků v lesních školkách). Na druhé straně snesou lesní dřeviny velice nízké teploty až -60 °C, podmínkou je však zpravidla předešlá příprava a stav dormance. Rostliny původem z nízkých zeměpisných šířek bývají poškozeny již při dosažení teploty mrazu.
Naše rostliny jsou přizpůsobené střídání denních a nočních teplot. Ale při přehřátí osluněného kmene za dne a následnému rychlému snížení teplot za noci může docházet k poškozování vnějších pletiv.
Průměrná roční teplota v naší republice je zhruba +7-8 °C (v horách +5 °C,v teplých nížinách +10 °C). Průměrná měsíční v nejchladnějším měsíci(lednu) - 2-3 °C, v nejteplejším měsíci (červenec) +18-20 °C.
Teplota má zásadní význam i pro intenzitu základních životních reakcí a projevů rostlin,
jako je transpirace, asimilace a respirace a její dynamika má primární vliv
na rozšíření vegetace a vznik vegetačních pásem na zemském povrchu.
Jako ekologicky významná je např. délka vegetační doby , označující
počet dnů, kdy je denní průměrná teplota alespoň 10 °C
(průměrná denní teplota je průměr 3 měření v průběhu dne (teplota v 7 hod. +teplota ve 14hod. a 2x teplota ve 21 hod./4). Pro existenci
lesního ekosystému je nutná její délka alespoň 1 měsíc, listnatý
opadavý les vyžaduje alespoň 4 měsíční trvání - to platí jak pro
zeměpisnou šířku, tak i pro nadmořskou výšku lokality.
Pro růst a
vývoj lesních dřevin jsou významné i extrémy, jako je například výskyt
pozdních a časných mrazů, na které jsou různé dřeviny různě citlivé.
Známá je třeba citlivost mladých buků na pozdní mrazy, limitující
obnovu bukem na holé ploše na celé řadě lokalit. Pionýrské dřeviny
se naopak vyznačují značnou tolerancí vůči klimatickým extrémům,
v první řadě pak extrémům teplotním.
Les a tepelné záření
Lesní prostředí modifikuje výrazným způsobem teplotní poměry lokality.
Na hodnoty teplot má kromě druhového složení a hustoty porostu vliv i jeho věk
a struktura.
Vliv porostu na průměrnou denní teplotu roste
v závislosti na jeho hustotě, čím hustší porost, tím nižší jsou v něm kladné teploty ale omezeny jsou i výkyvy teplot.
S dalším rozvolňováním opět
průměrné teploty půdy rostou v důsledku snížení zápoje a většího přístupu světla
(tepla) na půdní povrch.
Rozdíly oproti volné ploše se pohybují řádově v hodnotách jednotek až
desítek stupňů rozdílu (paseka/porost) maximálních teplot na povrchu půdy a několika jednotek
v 10-20 cm hloubce v půdě. V ročním průměru teplot je rozdíl mezi lesem a volnou plochou následující: letní teploty v průměru o 1-6 stupňů nižší, zimní teploty v průměru o půl až jeden stupeň vyšší, a výrazně se snižuje počet dnů s teplotními extrémy.
4.7 Voda

Voda je abiotickým faktorem, který ovlivňuje opět celou biotickou složku ekosystémů. Jako většinová součást živých organismů je nezbytnou podmínkou pro život. Je produkčním faktorem, jehož dostatek nebo nadbytek určuje produkční možnosti rostlin a následně potravní zdroje živočichů. Vlhkost půdy pak ovlivňuje podmínky pro půdní edafon a možnosti rozkladu organické hmoty a tím i rychlost cyklu živin. Voda ovlivňuje i abiotické část ekosystémů- chemické procesy v půdě a zvětrávání matečné horniny, charakter reliéfu krajiny, odstínění slunečního záření mračny v atmosféře.
Z hlediska hodnocení tohoto faktoru v ekosystému jsou důležité nejen celkové srážky za rok, ale především jejich rozložení v průběhu roku a v průběhu vegetační sezóny. Voda je limitujícím činitelem v biomech s celkově velmi nízkými srážkami (pouště, polopouště), ale i biomech s nerovnoměrným rozložením srážek v průběhu roku (stepi a savany, středomořské tvrdolisté lesy, některé subtropické lesy).
Rostlinné tělo, respektive jeho fyziologicky aktivní části, obsahují vysoká množství vody, zpravidla 80 - 95 % (listy, vodivá pletiva).
Voda je základním prostředím veškerých biochemických procesů v rostlině, je nezbytnou součástí struktury buňky a umožňuje zachování homeostáze důležité pro správný průběh metabolismu,
má význam pro pevnost rostlinných pletiv a je součástí procesu fotosyntézy,
je médiem umožňujícím transport živin z půdy, kde se ve formě rozpuštěných minerálních látek (iontů) dostávají ke kořenům, odkud jsou selektivně
jednotlivé ionty přijímány a s proudem vody transportovány v rostlině k fotosyntetickému aparátu.
V opačném směru dopravuje produkty fotosyntézy do větví, kmene a kořenů.
Každý pokles obsahu vody pod optimální hodnotu vyvolává metabolické poruchy s následným poklesem produkce.
Hlavní síly pro proudění vody proti gravitaci jsou kapilární síly a podtlak vyvolaný transpirací.
V biosféře se voda vyskytuje v různých formách. Největší podíl, zhruba 1360 mil. km3 (97,1 %) je ve formě slané mořské
vody světových moří a oceánů, zaujímajících 71 % zemského povrchu.
Obsahuje proměnlivá množství rozpuštěných minerálů,
z nichž převažuje Na+, Mg2+, Cl- a SO2-, zatímco v tzv. sladké vodě na pevninách (2,9 % objemu) převažují ionty Ca2+
a HCO3-.
Při průchodu biosférou a zvláště pak při průniku korunami stromů a svrchními horizonty půdy se v ní zvyšuje koncentrace nejrůznějších minerálních
prvků až na úroveň živného roztoku nezbytné ho pro existenci rostlin.
Zásoby sladké vody jsou ze 77 % tvořeny pevnou fází (ledovce a sněhová pokrývka) 22 % tvoří podzemní voda a pouze 1 %
představuje voda v jezerech, tocích a v půdě (van den Leeden et al. 1990). Toto relativně malé množství představuje vodu
využívanou suchozemskými ekosystémy, včetně všech ekosystémů lesních. V našich podmínkách je poměr jednotlivých frakcí odlišný,
půdní voda představuje asi 76 %, podzemní 18 % a povrchová voda v tocích a nádržích 6 % celkového objemu, zásoby představují
asi 36 % objemu srážek.
Takřka všechna voda na naše území spadá ve formě srážek, které jsou tak pro vodní bilanci rozhodujícím faktorem - z toho plyne i důležitá funkce lesů, zásadně ovlivňujících složky vodní bilance území. Množství vody v půdě je dáno množstvím dopadajících srážek, hloubkou hladiny podzemní vody, fyzikálními vlastnostmi půdy a rostlinstvem, pokrývajícím půdu. Obsah vody v půdě je výsledkem procesů vstupu vody do lesního ekosystému a ztrát vody z něj. Kromě bilance vody v ekosystému je pro jeho stav určující i charakter půd sledovaných lokalit. Půdní voda je hlavním a určujícím zdrojem vody pro rostliny včetně lesních dřevin. Její obsah a dynamika podmiňuje život rostlin přímo, jako zdroj vody pro transpiraci a s ní spojené transportní procesy v systému půda-rostlina, i nepřímo, prostřednictvím půdních mechanických, fyzikálních, chemických i pedobiologických vlastností. Jako zdroj vody je významná gravitační, kapilární, obalová a hygroskopická voda. Voda, kterou jsou rostliny schopny odčerpat z půdy, se označuje jako fyziologicky využitelná voda. Její množství závisí především na vlastnostech půdy, které určují její retenci, ale i sílu jejího poutání půdními částicemi.
Koloběh vody
Podmínkou rovnovážného stavu vody v ekosystémech je její koloběh. Kromě přísunu vody má zásadní význam pro pohyb a přesun látek v rozpuštěné i suspendované formě a je důležitý i pro usměrňování toku energie. Koloběh vody je poháněn sluneční energií - výpar z vodních povrchů, půdy i vegetace a vzdušné proudění, vítr a gravitační energií - pád a tok vody v kapalné formě i pád a posun ve formě pevné ve směru gravitačního spádu.
Velký koloběh vody (velký hydrologický cyklus)
Charakterizuje pohyb a transformace jednotlivých fází vody v globálním , planetárním měřítku. Počátečním bodem je přechod vody z kapalné fáze na fázi plynnou - výpar, evaporace, Na výpar se spotřebuje asi 25 % energie slunečního záření dopadajícího na zemský povrch. V tomto měřítku se jedná především o výpar z hladiny oceánů. Vodní pára je pak vzdušným prouděním přenášena i nad kontinenty a zde padá ve formě kapalný a pevných srážek, nebo kondenzuje na povrchu objektů. Ve směru gravitačního gradientu pak ve formě povrchového a podpovrchového odtoku směřuje zpět k oceánu, nebo se díky zásaku stává součástí podzemních vod. Ty mohou také vyvěrat a zúčastnit se povrchového odtoku. Část vody se opět vypaří, stává se součástí těl živých organizmů nebo tvoří zásoby půdní vody, mj. důležité pro život rostlin i živočichů.
Malý koloběh vody (malý hydrologický cyklus)
Charakterizuje pohyb vody, její dostupnost a formy v rámci plošně malého území, nebo v rámci konkrétního ekosystému. Probíhá nad pevninou, v krajinném měřítku a typem krajiny je výrazně ovlivňován. Jeho znalost umožňuje racionální využívání zdrojů vody pro nejrůznější účely, tj vlastní vodní hospodářství. Na úrovni lesního ekosystému je pro zásobu a využití zdrojů vody důležitá jeho vodní bilance, jež je výsledkem procesů souvisejících s hydrologickými cykly.
Vodní bilance
vztah mezi složkami příjmu, akumulace a výdeje vody pro určitý prostor v daném čase.
Množství a distribuce (rozdělení srážek) patří mezi základní klimatické (makro - a mezoklima) faktory prostředí.
Celkový roční úhrn srážek v našich podmínkách: 400 - 1200 (1400) mm. Jednotka - mm srážek - odkazuje na výšku vodního sloupce na ploše 1m2, 1 mm srážek tedy odpovídá objemu vody 1 litr na 1m2.
Rozložení srážek v ČR: (jaro 25 %, léto 40 %, podzim 20 %, zima 15%).
Příjmová složka vodní bilance
- Srážky vertikální a horizontální
- Přítok vody povrchový a podpovrchový
Výdajová složka vodní bilance
- Evaporace (výpar fyzikální)
- Intercepce
- Transpirace (výpar fyziologický)
- Odtok povrchový a podpovrchový
Vyjádření vodní bilance Z dlouhodobého hlediska je dáno rozdílem příjmové a výdajové složky, tedy porovnáním srážek a přítoku vody a odtoku a všech forem výparu vody. Z krátkodobého hlediska v rámci ekosystému je možné hodnotit přesněji, s připočtením bilance vody v půdě a tedy změnou vlhkosti půdy.
Srážky

- voda kapalného nebo pevného skupenství vyskytující se v atmosféře, na povrchu předmětů a Země.
Srážky představují rozhodující vstup vody do lesních aj. ekosystémů.
Lze je členit na kapalné a pevné, voda může ale do ekosystémů vstupovat i v plynné formě.
Na celkovém úhrnu srážek se sice rozhodující měrou podílí srážky vertikální (déšť, sníh),
za určitých podmínek mají větší význam rovněž srážky horizontální (usazené) (zkondenzovaná mlha, námraza). Kromě úhrnu srážek hraje významnou roli i jejich
intenzita a distribuce - rozložení během roku.
Vlastní dešťové srážky představují hlavní, rozhodující část srážkového úhrnu. Srážky jsou hodnoceny z hlediska intenzity, doby trvání
a plochy, kterou zasahují. Deště z tepla, konvekční srážky, vznikají při ochlazování vlhkého ohřátého vzduchu při jeho výstupu do
větších výšek. Jsou charakteristické relativně malou zasaženou plochou, krátkou dobou trvání a zpravidla vysokou intenzitou,
až přívalového charakteru.
Depozici pevných srážek (sníh, námraza, led) a rovněž jejich tání ovlivňuje nadzemní část vegetace. V lesích probíhá ve
srovnání s holou plochou odlišně jak proces depozice sněhu, tak jeho transformaci a odtávání. Čerstvý sníh bývá kyprý a jeho
hustota je velmi nízká, 0,02 - 0,27 (hustota vody je 1 g/cm3),proto je třeba brát v úvahu vysokou zhmotnost sněhové pokrývky, zvláště u vlhkého sněhu.. Hustota ulehnutého sněhu je pak 0,20 - 0,40 g/cm3.
Hustota sněhu je v lesích nižší, roste tak i jeho izolační schopnost a klesá promrzání lesní půdy ve srovnání s bezlesím.
Evaporace
- přechod vody z povrchu půdy, sněhu nebo ledu do plynného skupenství (fyzikální výpar).Vyjadřuje se výškou sloupce vody v mm.
V užším slova smyslu představuje výpar z povrchu půdy, která nemá žádný vegetační kryt, nebo z povrchu půdy pod vegetačním krytem.
Jedná se o neproduktivní výpar, který znamená ztrátu vody z ekosystému (půdní složky a posléze celkového ekosystému).
Úroveň evaporace ovlivňují především klimatické podmínky a půdní poměry: půdní vlhkost, intenzita vzlínání vody v půdě, reliéf
(sklon, expozice, nadmořská výška), hloubka hladiny spodní vody, teplota, vítr, vlhkost vzduchu. Jemná struktura půdy úhrnný výpar zvyšuje,
stejně jako tmavá barva. Na vodou nasycených půdách závisí výpar na pohybu vzduchu, dosahuje hodnoty až 10 - 15 mm za den.
V podmínkách lesních porostů je výše výparu z povrchu půdy silně ovlivněna krytem půdy vegetací, tj. složením, strukturou,
zakmeněním a zápojem porostu. Lesy mají relativně velice nízký podíl neproduktivního výparu, průměrně kolem 10 % z celkového výparu
(louky 25 %, pole 45, obnažená půda 100 %).
V praxi je někdy nesnadné odlišit transpiraci a evaporaci, proto se uvádí souhrnná hodnota evapotranspirace.
Odtok vody
U odtoku vody je rychlost závislá na překážkách , bránících proudění vody- tj. charakteru povrchu, na tom, jestli se jedná o tok neusměrněný, po vrstvě opadu, nebo bylinné vegetace, nebo tok usměrněný, v erozní rýze, korytě potoka- rozdíl rychlostí je pak až více než desetinásobný (rychlost proudění vody ve vodních tocích je ca 2-4 m/sec.). Po kontaktu s povrchem půdy se může část vody opětně vypařit (evaporace), ulpět na povrchu půdy a nebo pronikat do jejího nitra, do hlubších půdních vrstev (vsak, infiltrace). Intenzita vsaku závisí na půdním krytu, struktuře a textuře, vodivosti, pórovitosti, vlhkosti půdy, stejně tak na jejím mechanickém narušení. Vsak zrychluje prokořenění půdy a aktivita půdní fauny - vzniká velké množství drénů, kanálků, s vysokou schopností vedení vody. Schopnost vsaku je zvýšena především mocnými vrstvami nadložního humusu, který nejenže tlumí energii dešťových kapek, brání dezintegraci půdy a povrchové erozi
Je-li rychlost vsaku nižší než množství vody dopadající na povrch půdy, může se voda hromadit na povrchu a popřípadě odtékat
ve směru gravitačního spádů - vytváří se odtok, respektive povrchový odtok.
Voda pronikající půdou, gravitační voda, se pohybuje ve směru gravitačního spádu, vertikálně, dokud není ovlivněna hlubšími
nepropustným vrstvami a sklonem svahu. Potom se při snížené schopnosti pohybu do strany může hromadit, stagnovat, a podílet se na
specifických pedogenetických procesech, např. oglejení v důsledku zamokření. Je-li mezi zamokřenými horizonty povrchový humus, lze
pozorovat v takto vzniklých anaerobních podmínkách rašelinění.
Při vhodných poměrech se může voda pohybovat i v
horizontálním směru a vzniká tak podpovrchový odtok, který je řádově pomalejší ve srovnání s odtokem povrchovým a
nepředstavuje nebezpečí eroze.
Při překročení infiltračního potenciálu stéká voda po povrchu většinou nejprve ve
víceméně souvislé vrstvičce jako nesoustředěný povrchový odtok, později dochází k soustředěnému odtoku k v rámci o proudů, tj. rýh a vodních toků. Povrchový odtok
představuje značné ohrožení půdní dynamiky, a to především v antropogenně pozměněných podmínkách.
Lesní půda se na rozdíl od jiných ekosystémů výrazně a efektivně podílí na transformaci povrchového
odtoku na podpovrchový.

Odtok vody a lesní porosty
- zpravidla poněkud menší objem odtékající vody
- vyšší vyrovnanost odtoku
- výrazně nižší obsah suspendovaných látek
- výrazně nižší obsah rozpuštěných látek
Lesy ovlivňují jednak vstup vody do půdy, ale i její ztráty z ní. Intenzívní transpirace se podílí rozhodujícím způsobem na snižování obsahu půdní vody. Při vytěžení porostu má narušení této funkce za následek zamokřování půd na holinách, alespoň v prvních letech po likvidaci porostu, ve větší míře na uléhavých půdách. Tato skutečnost pak může ovlivnit charakter bylinné vegetace na těchto stanovištích. V důsledku změněných půdních a stanovištních vlastností (struktura, obsah humusu, výpar, odtok) na holinách pak po určité době může dojít naopak k výraznému zvýšení výsušnosti lokalit.
Intercepce

Charakterizuje zadržování srážek korunami stromů a následný výpar z povrchu vegetace
Intercepce (záchyt, zadržení) = rozdíl spadu srážek na volné ploše a pod porostem.
Intercepce je významná položka vodní bilance lesních porostů. Voda, jež ve formě především vertikálních srážek vypadá z atmosféry, se dostává nejdříve do kontaktu s vegetací.
Její část je na jejím povrchu zachycena a odpaří se zpět do atmosféry. Podílí se na ní nejen stromová, ale i keřová a bylinná
(travinná) vegetace a z hlediska srážek nejen kapalné, ale i pevné srážky (sníh, námraza - sublimace).
V absolutním množství je nejvýznamnější záchyt srážek kapalných.
Při dostatečné pokryvnosti je od počátku zejména méně intenzivní srážky zachycováno veškeré množství padající vody. Plné propouštění srážek do porostu nastává až po nasyceníintercepční kapacity povrchu vegetace, kdy povrch přestává poutat povrchovými vazbami vodu (skropná voda), další padající srážky pronikají na povrch půdy.
Transpirace
je fyziologický nebo tzv. produktivní výpar vody rostlinou
-výdej vodní páry z povrchu organismů ( difúzní proces)
Představuje nejvýznamnější složku výparu. Rostliny tak vypařují až kolem 98 % přijaté vody, zbytek zadržují v rostoucích pletivech. Transpirace představuje nejdůležitější položku výdeje vody lesním ekosystémem - až 60 %. V lučních ekosystémech se pohybuje její podíl až kolem 50 % a v polním 40 %. Její objem spoluurčují četné faktory vnější i vnitřní: sluneční záření, teplota, vlhkost půdy i vzduchu, vítr, imise apod. Ke vnitřním faktorům náleží druhové složení porostů, jejich věk, stáří transpirujících orgánů, obsah vody, struktura a tvar listu, povrchové struktury, orientace, stav kořenového systému aj.
Na poměru mezi příjmem vody kořenovým systémem a výdejem transpirací pak závisí vodní bilance rostliny. K výparu dochází již uvnitř listů a výdej páry je regulován otevíráním průduchů.
( Zahrnuje více než 80%, přes pokožku a kutikulu se odpaří jen 5-20% vody).
Jen 1- 5% vody,
která rostlinou proteče, je použita k metabolismu. Význam transpirace
spočívá především v ochlazování povrchu, dále má fyzikálně velký význam
pro příjem rozpuštěných živin spolu s
vodou přes kořeny.
Výdej vody funguje zejména přes průduchy ve formě vodní páry do ovzduší a podle typu se dělí na následující:
- Transpirace stomatární
- výdej vody štěrbinou průduchů. Může být díky strmému gradientu par rychlejší než výpar z vodní hladiny. Rostlinou je rychle regulovatelný. - Transpirace kutikulární
- kutinizovaným povrchem epidermis (30 % u vodních r., u jehličnanů 3-10 %). Z krátkodobého hlediska rostlinou neovlivnitelný, z dlouhodobého možnost přizpůsobení . - Transpirace rhizodermální
- výpar vody z kořenů. Tento výpar není rostlina schopna regulovat. - možné je i vylučování vody v kapalné formě, především v podmínkách s vysokou vzdušnou vlhkostí - gutace
tento způsob se více uplatňuje u rostlin vlhkých tropů, typickým příkladem našich rostlin, které tento způsob využívají je např. kontryhel
Transpirační koeficient - množství vody (l,t) spotřebované na transpiraci rostliny či porostu za vegetační období
na jednotku hmotnosti vytvořené sušiny (kg, t)
Ve srovnání s jinými rostlinami je u lesních dřevin relativně nižší.
Příklad obecně uváděných hodnot pro transpirační koeficient: polní plodiny 300-900, lesní dřeviny stinné 170-340, -slunné 260-340,
buk 169, smrk 231, borovice 300, bříza 317, dub 344). Spotřeba vody se výrazně mění s věkem porostu, v první řadě ve srážkově
chudších oblastech. U borovice je nejvyšší kolem 40. roku, u dubu a smrku kolem 60. roku věku.
Tvoří přitom 45 - 70 % srážkového úhrnu. Z našich lesních dřevin využívá vodní zdroje nejhospodárněji
pravděpodobně buk, naopak nejméně ekonomicky rychle rostoucí dřeviny (topoly, vrby, v jižnějších zemích eukalypty).
faktory ovlivňující transpiraci:
Vnitřní: charakter rostliny, její možnosti hospodaření s vodou, fenologická (vývojová) fáze, stáří, biomasa. Vnější - teplota, pohyb okolního vzduchu, vzdušná vlhkost, tvar povrchu,
ze kterého se voda odpařuje
4.8 Půda

Půda je významnou složkou lesních ekosystémů, která má rozhodující vliv na dostupnost živin a tím i na produkční možnosti daného ekosystému. Je zásadním prostředím, přes které probíhá koloběh živin a tok energie, prostředím pro život půdních organismů (reducentů) jako koncových částí potravních řetězců, půda a její charakter určuje vzhled terénu, možnosti pro růst rostlin a úkryt živočichů, čímž ovlivňuje všechny části biotické složky ekosystému. Z abiotických složek ovlivňuje vodní režim - výpar a možnosti zásaku a povrchového odtoku vody a tím i dostupnost vody rostlinám a živočichům.
Složení půd z fyzikálního hlediska

jako taková, jako stabilní prostředí zemského povrchu,
je tvořená souborem různých prostředí a látek, které se navzájem prolínají, ovlivňují a pozměňují
základním rozdělením hmoty tvořící půdu je rozdělení na pevnou, kapalnou a plynnou fázi.
Plynnou fázi tvoří půdní vzduch s vyšším obsahem oxidu uhličitého a nižším obsahem kyslíku, než vzduch atmosférický. Půdní voda- půdní roztok má také vyšší koncentrace různých rozpuštěných iontů než srážková voda, a je zásadním prostředím pro průběh velké části chemických reakcí v půdě.
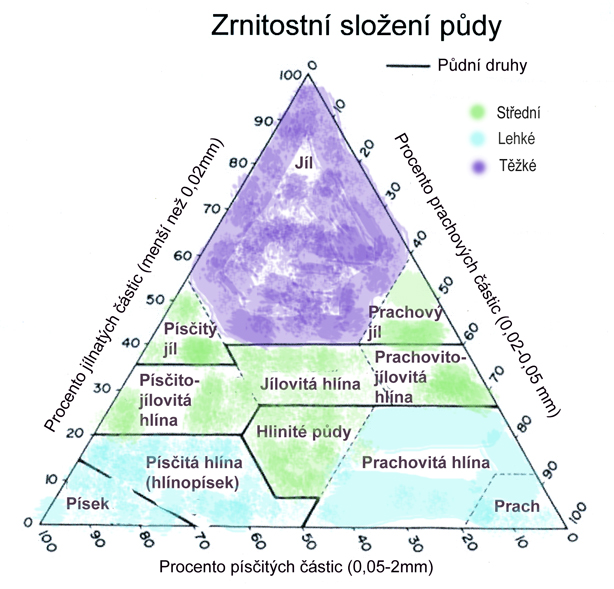
Vzájemný poměr těchto půdních složek pak určuje řadu jejích fyzikálních vlastností (měrnou hmotnost, schopnost poutat vodu, propustnost, vysýchavost, vedení tepla v půdě, objemové změny při promrzání...). Základem pro tyto fyzikální vlastnosti je zrnitostní složení pevné fáze. Jednotlivá zrna na sebe těsně nepřiléhají a podle své velikosti vytvářejí v půdě póry (hrubé, jemné, kapilární, nekapilární), které mohuo být zaplněné vodou nebo vzduchem. Světlost pórů pak určujě, jakým způsobem se v půdě bude pohybovat kapalná fáze (možnosti vzlínání, rychlost zásaku).
Podle zrnitostního složení (textury) určujeme- Lehké půdy
- Středně těžké půdy
- Těžké půdy
Složení půd z chemicko-ekologického hlediska
- Minerální složka půdního prostředí 93%
- Organická složka půdního prostředí 7%
- Živá složka: (biomasa)
- Kořeny rostlin 10%
- Půdní organismy (edafon)5%
- Žížaly 12%
- Ostatní makrofauna 5%
- Mikro a mezofauna 3%
- Bakterie, aktinomycety 40%
- Houby, řasy 40%
- Neživá složka: (nekromasa)
85% odumřelá organická hmota v různém stádiu rozkladu
- Živá složka: (biomasa)
Minerální složka půdního prostředí
Pedochemie jednotlivých prvků (jejich zastoupení, formy výskytu, typy vazeb, chemické reakce, zapojování do biologických cyklů apod. těsně souvisí s jejich postavením v Periodické tabulce prvků (poloměr a hydratace iontů, valence) a s jejich zastoupením v horninách a s jejich geochemickým a biochemickým chováním.
Minerální složka půdního prostředí je dána především matečnou horninou, na povrchu které se půda nachází a obsahuje tudíž její části - jednotlivé minerály tuto horninu tvořící a dále produkty rozpadu a zvětrávání těchto minerálů.
- zrna minerálů obsažených v matečné hornině, chemicky nezměněné zbytky minerálů s obtížnou zvětratelností, jako je např. křemen, zirkon, turmalín...
- sekundární minerály (především jílové minerály, hydroxidy, silikáty.....), vzniklé v půdě v důsledku chemických přeměn a transportu iontů v půdním roztoku. Některé sekundární minerály mohou tvořit půdní koloidní minerální komplex schopný poutat a uvolňovat ionty do půdního roztoku.
- Soli, oxidy, hydroxidy uvolněné do půdy v rámci chemického zvětrávání matečné horniny a schopné dále reagovat, podílet se na výměně iontů a chemismu půdy.
Zastoupení jednotlivých prvků v půdách a jejich vlastnosti
- Kyslík - nejrozšířenější, ve více než polovině známých minerálů v půdách dále složkou vody, vzduchu a organických sloučenin
- Křemík - křemen a silikátové a alumosilikátové minerály v křemičitanech centrálním iontem SiO4tetraedrů, menší množství v amorfní kyselině křemičité, opálu vázaný v silikátech (nerozpustný 20% HCl) je pro půdu chemicky neaktivní v alumosilikátech (jejich zvětratelný podíl je v HCl rozpustný) má význam v chemické dynamice půdy (koloidní kys. křemičitá a křemičitany) v půdách sorpčně nenasycených e koloidní kyselina křemičitá a silikáty pohyblivé ve formě hydrosolů v půdách sorpčně nasycených jsou koloidní křemičitany nepohyblivé
- Hliník - silikáty, oxidy, hydroxidy, centrálním iontem Al-tetraedrů a Al-oktaedrů. Přechod Al z tetraedrické do oktaedrické konformace (při zvětrávání) a opačně (při metamorfóze) je z hlediska geochemie poměrně významnou reakcí a vede k uvolnění Al, jeho rychlé hydrataci Al (H2O)6 a tvorbě polymorfních hydroxidů. Chemismus v půdách je dán jeho amfoterní povahou, snadnou hydrolyzovateností a polymerací v půdách sorpčně nasycených jsou koloidní sloučeniny hliníku nepohyblivé jako hydrogely v půdách sorpčně nenasycených pohyblivé ve formě hydrosolů
- Železo - tvoří oxidy, sulfidy, fosfáty, silikáty Při zvětrávání oxidováno, vzniká červenohnědé zbarvení, v anaerobních podmínkách v redukovaném stavu šedavé až nazelenalé zbarvení půdy. V půdách většinou sloučeniny ve formě koloidního hydrosolu (pohyblivé v půdním profilu), jejich pohyblivost se zvyšuje působením kyselého humusu.
- Uhlík - karbonáty, organické sloučeniny, oxid uhličitý v půdě asi 20x víc než v litosféře
- Vápník - plagioklasy, pyroxeny, amfiboly, kalcit, dolomit, anhydrid, sádrovec, fosfáty, fluorit má významnou úlohu v udržení pH, pufračních schopnostech půd, tvorbě půdní struktury (působí koagulaci koloidů), zvyšuje přístupnost některých živin, snižuje aktivitu volného hliníku
- Hořčík - podobné vlastnosti, snadno se vyluhuje z půd, nebo se při výparném režimu hromadí ve svrchní vrstvě půdy
- Draslík - silikáty (ortoklas), slídy, halogenidy snadno pohyblivý v půdě, ale o něco hůře vyluhovatelný než sodík
- Dusík - podstatnou složkou humusových látek obsah v půdě 0,1-0,3%
- Fosfor - apatit, částečně silikáty (fosforečnanový iont substituovaný v Si tetraedrech, organické sloučeniny
v půdách s pH nad 7,2 převládají Ca, Mg fosfátové komplexy
při pH 4,3-7 hydrolyzovatelné fosforečnanové anionty
při pH pod 4,3 Al, případně Fe fosfátové komplexy - Mangan- dvojmocný i trojmocný, v kyselém prostředí pohyblivější než železo; při vyšším pH může tvořit konkrece s vysokou sorpční kapacitou (pro Co, Cu, Ni)
- Síra - sulfidy, sírany, organická hmoty
- Vodík - součástí asi 1/3 minerálů, kde vytváří hydroxylové skupiny v krystalové mřížce, dále vody, organická hmota
- Titan - oxidy a silikáty, v půdě málo pohyblivý
- Bór - turmalín, aniont B(OH)4- adsorbován hydroslídami, v pedosféře 3x více než v litosféře
- Chrom - při zvětrávání se hromadí v jílovém podílu
- Kobalt - při zvětrávání se uvolňuje a hromadí spolu s manganem v půdních novotvarech
- Nikl - součástí Fe a Mg minerálů
- Molybden - může se hromadit v jílu a humusu, výskyt ve formě 3 a 6mocného, v mřížkách jílových minerálů může nahradit Al a Fe.
- Měď - součástí sulfitů, zvýšená pohyblivost v komplexu s humusovými kyselinami
- Zinek - součástí sulfitů, absorbován v komplexu s jílem a hydráty sesquioxidů
Organický podíl v půdě
Dynamická složka, ve které dochází k rychlým tokům látek a energie, a která určuje charakter vývoje půd
Živá část organického podílu- půdní organismy
účastní se v půdě většiny pochodů přeměn organické (někdy i minerální) hmoty procesy probíhají buďto v tělech organismů, nebo působením enzymů mimo jejich těla. Po odumření mohou také dodávat organickou hmotyu do půdy.
Kořeny rostlin

Kořeny rostlin se vyskytují především v povrchové vrstě - zvláště pak jemné kořeny, ve svrchních 15-20 cm. Jejich množství závisí na typu rostlinného pokryvu - u lesních porostů mírného pásu tvoří kořeny ca 10-20% biomasy rostlin, u tropických deštných lesů s velkým množstvím epifytů vca 7-10% nadzemní biomasy, u travních společenstev může být biomasa kořenů větší, než biomasa nadzemní části rostlin.
Mikroedafon
- množstvím i hmotností je to převládající složka. Požadavky na prostředí jsou u různých skupin různé: vzhledem k tomu, že často jsou skupiny navzájem potravními konkurenty, mohou být i protichůdné. Přesto pro vysokou biodiverzitu mikroedafonu je zapotřebí dostatečná vlhkost a zároveň i provzdušnění půdy, dostatečná teplota a zásoba živin a mrtvé organické hmoty v půdě.
- Bakterie
s množstvím 10 na šestou (106) až 10 na devátou v 1g půdy jsou skupinou nejpočetnější. Druhy aerobní, fakultativně anaerobní, anaerobní
- podíly a druhové zastoupení závisí na vodním a vzdušném režimu, zásobení živinami, vegetačním krytu
Různorodá biochemická funkce:
Rozklad cukrů, bílkovin, celulózy, částečně i ligninu
Přeměna org. látek: humifikace, oxidace amoniaku, fixace vzdušného N, oxidace Fe, S, Mn, rozklad síranů, dusičnanů
rody: Pseudomonas, Bacillus, Micrococcus, Achromobacter, Clostridium, Arthrobacter, fixátoři: Azotobacter, Rhizobium, Clostridium,...
přeměny dusíku: Nitrosomonas, Nitrobacter - Houby
účastní se na procesech rozkladu tuků, bílkovin, uhlovodíků, ligninu
vyšší houby symbionty kořenů rostlin (mykorhiza)
nejčastější skupiny: Mucor, Penicillium, Trichoderma, Aspergillus...
- Aktinomycety
citlivé na reakci prostředí (neutrál až alkalické),vlhkost a teplotu
podílejí se na rozkladu organ. látek často pro jiné druhy rezistentních (lignin, chitin, humusové látky,...)
skupiny: Streptomyces, Nocardia, Actinomyces, Micromonospora ... - Řasy a prvoci
fotoautotrofní, v povrchových vrstvách půdy, kladně reagují na vlhkost a živiny
hlavní funkcí je hromadění organické hmoty ve vlastních tělech
skupiny: sinice Cyanophycae, rozsivky Diatomae, zelené řasy Chloropyceae ...
Prvoci jsou většinou heterotrofní, zřídka autotrofní, převážně aerobní
živí se bakteriemi, jejichž počet mohou při svém přemnožení (pařeniště, skleníky) redukovat
skupiny: nálevníci Cilliata, bičíkovci Flagellata, kořenonožci Rhizopoda
Mezoedafon
Typickými zástupci jsou například: háďátka, chvostoskoci, žížaly, hlísti a hlístice, stonožky, mnohonožky, roztoči, hmyz a jeho vývojová stádia, měkkýši
Patří sem zástupci živočišné říše, mimo obratlovců.- geobionti
- tráví celý život v půdě - geophila
- v půdě pouze vývojová stádia, dospělci nikoli - geoxena
- v půdě víceméně náhodně
Vyžadují dostatečnou zásobu organických zbytků, dobré provzdušnění, příznivé pH, vlhkost,
teplotu.
Výskyt je především v povrchové vrstvě 10-15cm, menší část do 35 cm (žížaly) ojediněle až do 1m.
Hlavní význam spočívá v mechanickém kypření a rozrušování půdní hmoty, vytváření chodbiček a kanálků (provzdušnění
a průnik vlhkosti). Dále mechanické rozrušování a přesun organických zbytků. Dešťovky a nižší červi krom toho
ještě obohacují výměšky o minerální částice a uhličitan vápenatý. Vytvářejí pevné a vodostálé agregáty.
Makroedafon
Typickými zástupci jsou: krtci, sysli, křečci, hraboši a další myšovití
Vliv makroedafonu na půdní prostředí je spíše omezený. Hlavní funkce spočívá zejména v mechanickém prohrabování,
provrtávání půdy a přesunu (zatahování) organické hmoty do vyšších hloubek
Neživá organická hmota
Zbytky rostlinných a živočišných organismů v různém stupni přeměny, nacházející se v půdě, nebo na jejím povrchu.
Význam organické hmoty v půdě spočívá především v zachování koloběhu živin
- návrat živin uložených v biomase zpět do půdy.
Organická hmota má na půdu i další vliv
- ovlivňuje její schopnost poutat živiny, tím, že zvyšuje množství půdních koloidů a brání tak jejich ztrátě
z ekosystému např. vyplavením.
Dále zlepšuje vlastnosti půdy jako je tepelná vodivost, provzdušněnost,
schopnost poutat vodu, snižuje výpar a promrzání, zlepšuje její strukturu.
Na tuto organickou hmotu lze pohlížet z různých hledisek a podle toho ji dále dělit, nebo hodnotit:
Podle stupně přeměny:
- celkový humus
- veškerá organická hmota v půdě, nekromasa, bez ohledu na původ, stav rozkladu a její umístění v rámci půdy - vlastní humus
- organická půdní hmota která prošla kompletně humifikačními transformačními procesy(rozklad, kondenzace, polymerace, tvorba aromatických sloučenin) procesy díky činností půdních organismů a jejich enzymů.
Z chemického hlediska jde o soubor tmavě zbarvených organických dusíkatých polyfunkčních látek vysoké molekulové hmotnosti, kyselinové povahy, převážně koloidního charakteru (Tyto humusové koloidy jsou schopné na svém povrchu poutat a tím i v půdě uchovávat živiny), relativně odolných vůči mikrobiálnímu rozkladu. Dále je typická cyklická struktura jádra, vazba na minerální půdní podíl a amfoterní charakter.
Podle chemického složení:
- nespecifické humusové látky
(nehuminové a primární)
organické kyseliny, glycidy, pektiny, bílkoviny, třísloviny, tuky, vosky, pryskyřice
tvoří energetickou a živinnou zásobu půdy a jsou podmínkou biologické aktivity půdy - specifické humusové látky
(huminové a sekundární)
vysokomolekulární organické sloučeniny, tvoří 85-90% org. hmoty v půdě
Podle funkcí v půdě:
- materiál humusotvorný
(organická hmota nerozložená) - meziprodukty humifikačních procesů
- vlastní humus jako výsledek humifikace
Podle uložení v půdě a stupně rozkladu:
Nadložní humus
Horizont L
- opad - různě starý opad listů, jehlic, částí dřeva, které většinou nejsou ještě mechanicky výrazně poškozené, ale už na nich jsou patrné barevné změny (vyluhováním)Horizont F
- vrstva mechanicky rozdrobené organické hmoty, částice velikosti několika až 1 mm, rozpoznatelné částice převažují nad jemnou frakcí, chemické změny díky intenzivní činnosti půdních organismů - žížal, hub, bakterií, rovněž je zde značný výskyt jemných kořenů rostlinHorizont H
- rozložená organická hmota, na jejíchž částečkách již není poznat jejich původ, převažuje jemná frakce, vyjímkou mohou být větší části kůry a dřevaSvrchní půdní horizont
- horizont Ah , jež obsahuje minerální zrna a organický podíl (do 30%)
Podle formy
Podle typu uložení jednotlivých vrstev, které různě rozložená organická hmota vytváří a podle způsobu a intenzity rozkladu organické hmoty na různých stanovištích v různých podmínkách se odlišují tři základní humusové formy. Tato klasifikace pomáhá určit typ a bohatost stanoviště, rychlost koloběhu živin v daném prostředí a možnost odhadnout možnou biodiverzitu a často i zastoupení jednotlivých druhů rostlin.
Mull
Vznik na bohatých většinou dostatečně vlhkých a provzdušněných černozemích a nivních půdách.
Horizonty L, F, H mají menší mocnost než horizont Ah a ta se mění v průběhu roku.
Silná humusotvorná činnost, humifikační procesy vysokého stupně, převaha huminových kyselin, vysoká kationtová výměnná kapacita...
Velká biodiverzita půdních organismů, vysoké zastoupení žížal (50-80%), členovci, mnohonožky, brouci, z mikroflóry zejména bakterie, aktinomycety.Moder
Nejrozšířenější forma na hnědých lesních půdách, vznik většinou na kambizemích, středně bohatých půdách.
Horizonty L, F, H mají větší mocnost než Ah (Ah do 10 cm), horizont F je zoogenní, nebo přechodný.
Střední intenzita humifikačních procesů, slabá syntéza humusových komplexů, vyšší podíl fulvokyselin než u formy mull.
Méně žížal, členovci, roztoči, chvostoskoci, z mikroflóry jsou zde aktinomycety, houby, bakterie...Mor
Vznik na chudých půdách s nedostatkem živin, často suchá nebo kyselá stanoviště, písčité půdy, podzoly Vysoká mocnost nadložního humusu, horizonty L, F, H výrazně přesahují mocnost Ah, ta je menší než 2 cm, horizont F vrstevnatý, vzniklý činností hub.
Pomalé rozkladné procesy, kyselý povrchový humus, nízká KVK, fulvokyseliny.
Vznik humusu především činností hub, což se projevuje vrstevnatou strukturou horizontu F.
Z půdních organismů hlavně roztoči, chvostoskoci, z mikroflóry zejména houby, méně aktinomycet.
Vlastní humus
lze dále rozdělit na několik skupin látek, podle intenzity a stupně humifikačních procesů, které zde proběhly:
-
Humínové kyseliny
Jsou nejhodnotnějším produktem humifikačních procesů v půdě, ovlivňují možnou vazbu kationtů, strukturu a vysokou pufrační schopnost půdy- základní složkou fenolické jádro, jsou to cyklické dusíkaté sloučeniny
- molekulová hmotnost 300 000
- C- 52-62%, H - 2,8-5,8%, 0- 31-39%, N 1,7-4,9%, 1-10% popelovin (Si, Al, Fe, S, P, Ca, Mg, K, Na)
- tmavá barva, rozpustnost v hydroxidech a roztocích zásaditých solí, snadná srážlivost dvojmocnými kationty, špatná rozpustnost ve vodě
- v nasyceném stavu stálé, odolné proti mineralizaci
- Kyselinový charakter daný přítomností -COOH skupin, a fenol-OH
- schopnost vyměňovat vodík, v neutrálním prostředí 250-500 mmol/100g
- porézní stavba, vysoká sorpční schopnost (400-600 mmol/100g)
součást humínových kyselin, lze je oddělit extrakcí alkoholem mají žlutohnědé zbarvení a menší molekulovou hmotnost Fulvokyseliny
v důsledku silně kyselé (pH 2,6-2,8) reakce jsou agresivní k minerální části půdy, kterou mohou ochuzovat o živiny a minerální látky- molekulová hmotnost je 200 - 50 000
- C - 40-52%, H 4-6%, O 40-48%, N 2-6% , popeloviny 2-8%
- jsou žluté až hnědé barvy
- velmi pohyblivé a lehce se přemísťují v půdním profilu
- dobrá rozpustnost ve vodě, minerálních kyselinách i zásadách, roztocích hydrolytických zásaditých solí výměnný vodík karboxylových skupin 600-900 mmol/100g
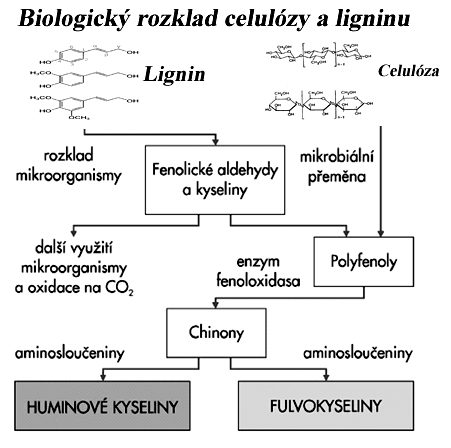
Humíny a humusové uhlí
- humíny někdy charakterizovány jako nerozpustná forma humínových kyselin
silně karbonizovaná organická hmota, pevně vázaná na minerální podíl půdy - humusové uhlí se vyskytuje v půdním humusu jako nejstarší složka produktů humifikace
tmavá, zuhelnatělá, na uhlík a dusík bohatá hmota, která nepeptizuje, nerozpouští se, nehydrolyzuje, nezúčastňuje se půdotvorného procesu
- humíny někdy charakterizovány jako nerozpustná forma humínových kyselin
Tvorba humusu
-
Mineralizace
V podmínkách příznivých teplot, vlhkosti, provzdušnění.
V půdě je v těchto podmínkách pozorovatelná silná činnost aerobních bakterií, které rozkládají organickou hmotu na jednoduché složky (CO2, H2O, NH3, oxidy)
v tomto případě se humus netvoří, nebo je velmi rychle rozkládán -
Rašelinění nebo uhelnatění
V podmínkách kyselé reakce prostředí, nízké teploty, vysoké vlhkosti, za omezeného přístupu vzduchu, anaerobních podmínek, nedostatečné oxidace
Nedokonalá přeměna organické hmoty - nedokonalý rozklad organický zbytků, enzymatický a biochemický proces způsobený zejména anaerobními bakteriemi
Výsledkem jsou huminové a ulminové látky tmavohnědé-černé barvy s vysokým obsahem uhlíku Humifikace
Optimální podmínky jsou periodické ovlhčování a vysýchání, střídání aerobních a anaerobních procesů.
Proces humifikace je převážně anaerobní, soubor převážně enzymatických a biochemických pochodů, při nichž se z meziproduktů rozkladu tvoří resyntézou tvoří tzv. huminové látky.
Mají obsah uhlíku k dusíku 1:10, hnědou až černohnědou barvu a vlastnosti koloidů.
Humifikace je provázena mineralizací výchozích látek (nikdy nezhumifikuje všechna org. hmota).
Článkem tvorby humusových látek je kondenzace strukturních jednotek, ke které dochází okysličením fenolů přes fermenty typu fenoloxidáz vedoucím k tvorbě chinonů a vzájemnou reakcí chinonů s aminokyselinami a peptidy, závěrečnou fází je polymerace (polykondenzace). Ta je reverzibilním procesem a proto je nutné aby byl produkt z prostředí odváděn, jinak může dojít k rozpadu již vytvořených kondenzátů (např. při nadbytku vody v půdě).
Vzájemný vztah mezi rostlinami a půdou
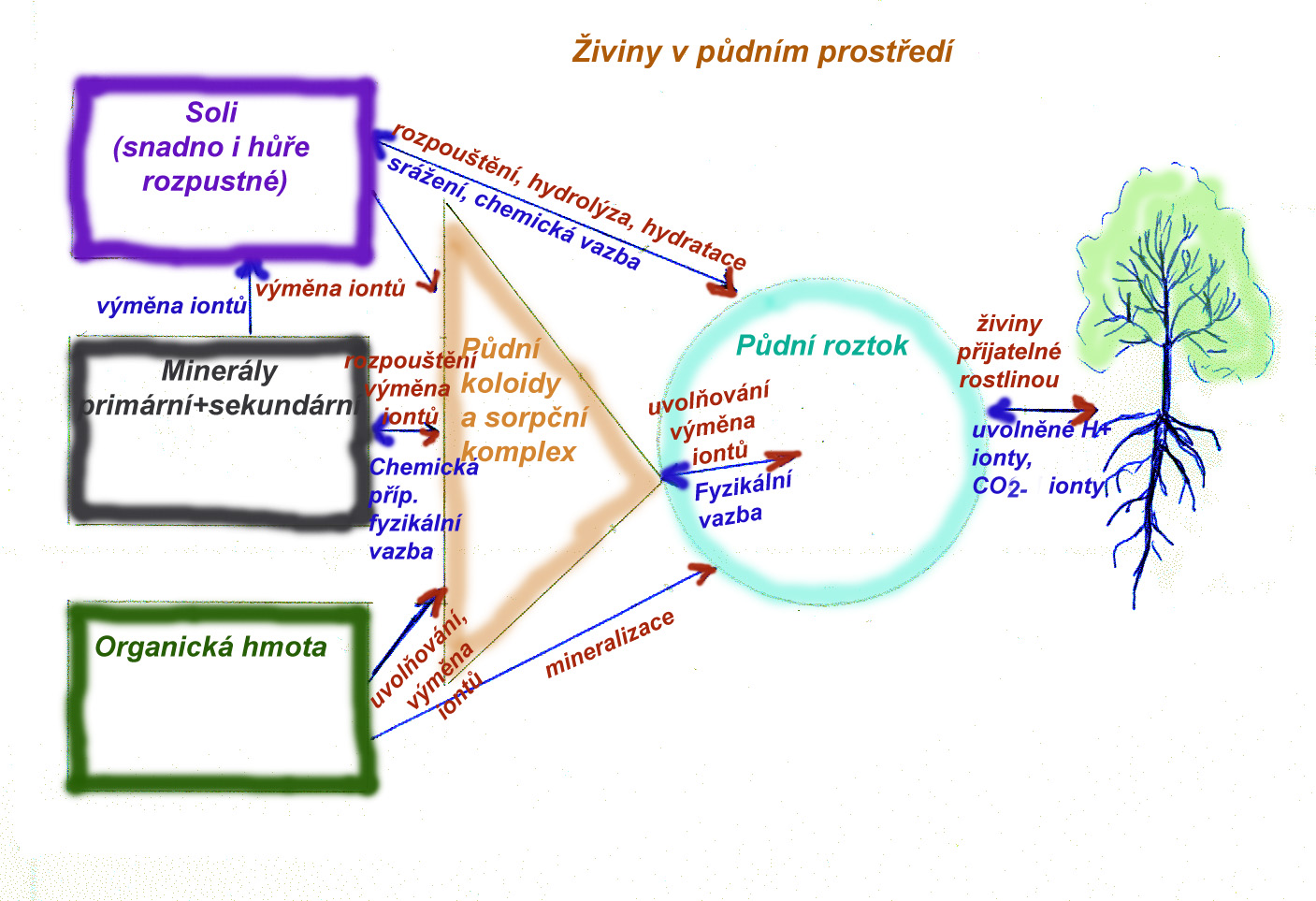
Akumulace organických látek v půdě je určena přísunem organické hmoty, jejich rozložitelností, klimatickými faktory, trofickými podmínkami pro
rozkladnou činnost mikroorganismů.
Každý typ vegetace je charakterizován rozložením nadzemní a podzemní biomasy, čistou primární produkcí, ročním opadem.
Odběr živin rostlinami, především pak dřevinnou vegetací, má zásadní význam na stav lesních půd.
Živiny jsou v půdě obsaženy v různé míře, tato úroveň je kromě jiných faktorů označována jako úrodnost půdy.
Po odběru jsou v půdním prostředí podle možností a podmínek jednotlivých stanovišť živiny nahrazovány
z následujících zdrojů (podle významnosti):
- mineralizace organické hmoty a zpřístupnění v ní obsažených živin
- zvětrávání matečné horniny a uvolňování živin obsažených v minerálech
- spad živin s atmosférickými srážkami (mokrá depozice) nebo suchá depozice prašných součástí (i antropogenních)
- vstup s vodou podpovrchovým nebo povrchovým tokem ze sousedství
- zvláštním případem je poutání (fixace) vzdušného dusíku
Při příjmu většiny živin rostlinami dochází k jejich nahrazování v půdním prostředí ionty s podobným nábojem,
což v případě kationtů znamená především ionty vodíku. Rovněž příjem aniontů, např. nitrátů, znamená většinou uvolnění vodíkových kationtů do půdního roztoku či sorpčního komplexu a tím i změny půdního chemismu.
V důsledku odběr živin rostlinami znamená acidifikaci půd a jejich vyčerpání ve vztahu k živinám.
Je-li vstup živin do ekosystému menší než jejich odběr vegetací, dochází k poklesu jejich obsahu v půdě, tj. k její degradaci. Závažnost tohoto jevu závisí na velikosti a rychlosti poklesu obsahu živin v půdním prostředí.
V lesích přírodních jsou většinou ztráty živin z půdy nahrazovány v dostatečném množství dekompozicí organické hmoty
a částečně dalším zvětráváním, v hospodářsky využívaných lesích je však ochuzení ekosystému často irreverzibilní pro
velký rozsah odběru a pomalé doplňování zásoby živin z přírodních zdrojů. Při větším odběru živin dochází k poruchám
látkových koloběhů a degradaci ekosystémů včetně snížení jejich produkce.
Návrat živin do půdy je daný z velké části kvalitou opadu, klimatickými podmínkami a jejich vlivem na činnost rozkládajících půdních organismů.
Kvalita opadu má určující význam pro celou řadu půdních procesů vedoucích k vytváření humusových forem. Určována je především obsahem živin, který determinuje i jejich potenciální vhodnost jako půdotvorného substrátu a substrátu pro vznik nadložního i vlastního půdního humusu. Rozložitelnost opadu dřevin je závislá na různých vnitřních faktorech,jako je např. obsah taninů a pryskyřic, impregnačních látek, částečně i živin a asimilátů, nicméně orientačně je možné použít především poměr C:N v tomto opadu.
Vznik půd
Vznik půd je složitý proces ovlivňovaný prakticky všemi složkami prostředí- průběhem klimatických faktorů, dostupností vody, charakterem matečné horniny, přítom,nými živými organismy a opadem rostlin. V průběhu tvorby půd dochází ke změnám fyzikálním (změna textury, promíchávání organického a anorganického materiálu) i změnám chemickým (vznik sekundárních minerálů, rozpouštění, srážení, přesuny iontů, tvorba a rozpad sloučenin).V průběhu vzniku půdy se postupně diferencují její jednotlivé horizonty, které následně mají rozdílné zrnitostní, chemické i funkční charakteristiky.
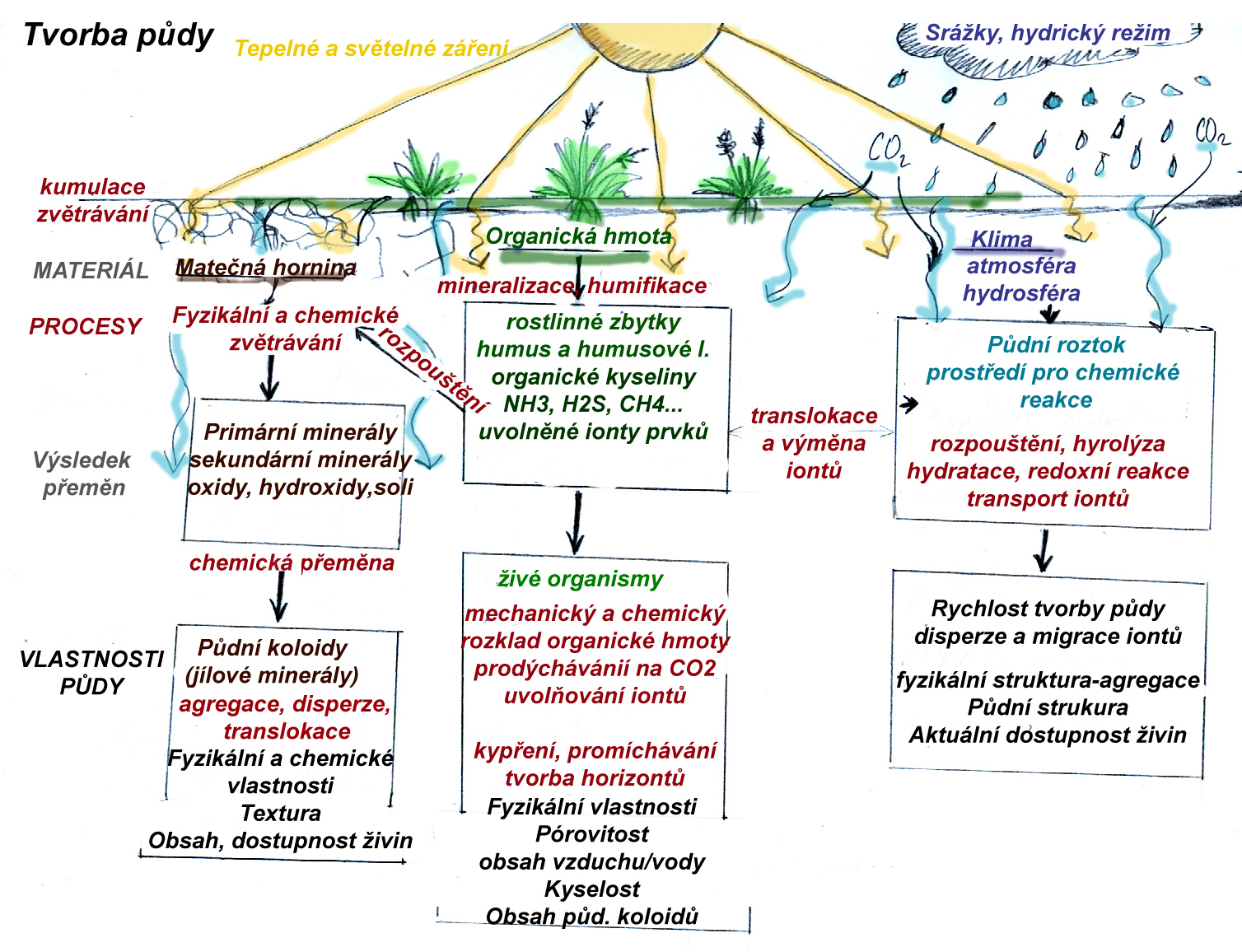
- Půdní horizonty:
- L+F+H- horizonty nadložního humusu
- A - svrchní minerální horizont, obohacený humusem (ca 20-30%)
- E- eluviální, vybělený/vymytý horizont, na přechodu mezi horizontem A a B (není vždy přítomný)
- B - minerální horizont (obsahuje jak primární, tak sekundární minerály, také malé množství organické hmoty. Dochází v něm k chemickýcm přeměnám - např.transportu iontů nebo jílnatých částic, díky kterým může být rozčleněn do více podhorizontů (Bh- obohacený humusem, B)
- C - půdotvorný substrát (mechanicky rozpadlý geologický substrát, tzv. skelet- může být totožnáý s matečnou horninou, nebo může jít o substrát mechanicky přemístěný (náplavy)
- R - matečná hornina (prakticky jen velmi slabě mechanicky navětraná)
Podle charakteru půdních horizontů je možné rozlišit jednotlivé půdní typy .
Půdní typy
Stručný přehled půdních typů, který je přesněji specifikován v detailních pedologických systémech, např.: http://klasifikace.pedologie.czu.cz/index.php?action=showKlasifikacniSystem
- Leptosoly - půdy s vysokou skeletovitostí a mělkým profilem
- Litozem- slabě vyvinutá, do 10 cm
- Ranker- více než 50% skeletu, na sutích,v pahorkatinách a hornatinách
- Rendzina- rozpad karbonátových hornin (vápence a dolomity),
- Pararendzina- z méně pevných karbonátových hornin, přechod ke kambizemím
- Regosoly- nevyvinuté půdy na chudých sypkých substrátech na rovinách (pouze horizonty A a C))
- Fluvisoly- fluvizemě, jsou půdy vznikající v nivách řek z povodňových sedimentů, významným znakem je vrstevnatost a nepravidelné rozmístění organické hmoty (0,5%) v celém profilu. Další variantou jsou Koluvisoly, rovněž vrstevnaté, vznikající hromaděním materiálů ve spodní části svahů.
- Černosoly- půdy s hlubokým humusovým horizontem (40-60cm), na bohatých horninách- karbonáty, apod.
- Černozem- na spraších a karbonátových slínech
- Černice- na těžších substrátech, vlhčích podmínkách, vyšší obsah humusu
- Vertisoly - půdy vznikající na jílech v suchých oblastech, typický je vznik trhlin za sucha: Smonice- půda vzniklá ze smektitických jílů
- Luvisoly - půdy obohacené o humus i v hlubších horizontech (horizont Bh, Ah), mají vyluhovaný E horizont
- Šedozem-obsahuje vybělený horizont ochuzený o jíl a tmavý luvický horizont B
- Hnědozem- mírně vybělený horizont E,stejnoměrně hnědý luvický horizont
- Luvizem- výrazně vybělený lístkovitý horizont E, ovlivněné vodou (promyvným režimem)
- Kambisoly- půdy s výrazným hnědnutím (braunifikací) vznikající na přemístěných zvětralých materiálech
- Kambizem- typický je kambický horizont, mají celkově velkou variabilitu
- Pelozem- mají pelický horizont s obsahem jílu nad 35%, vznikají na jílovitě zvětrávajících břidlicích
- Podzosoly- půdy kyselé, silně sorpčně nenasycené, s vysokým obsahem volného hliníku a železa
- Podzol- má vybělený Ep horizont, rezavě zbarvené znaky o pohybu železa, nízký obsah živin, Vznik buďto na píscích nižších poloh, nebo chudších horninách vyšších poloh.
- Kryptopodzol- má kyprý rezavě zbarvený horizot,
- Stagnosoly- půdy v oblastech s výraznějšímim srážkami, typické výskytem mramorovaného redukčního horizontu a vyšším obsahem amorfního železa v redukované formě
- Pseudoglej- mají výrazně mramorovaný horizont,na usazeninách i vybělený horizont a vysoký obsah humusu v ovrchových vrstvách
- Stagnoglej- vzniká při velmi dlouhodobém zamokření, má šedý glejový horizont
- Glejsoly- Glej- půdy vznikající při dlouhodobém provlhčení srážkami i podpovrchovou vodou, typické redukčním glejovým horizontem a rašeliněním organické hmoty v povrchových vrstvách
- Organosoly- půdy s rašelinnými horizonty nad 50cm, nad víceméně pevnou skálou
4.9 Atmosféra
Vítr- proudění vzdušných mas- je abiotickým faktorem, který má ve většině případů relativně menší přímý vliv na biosféru, avšak v některých případech může ekosystém ovlivnit poměrně zásadním způsobem. Je to faktor, který vzniká díky slunečnímu záření a rozdílné teplotě vzdušných mas.
Je to i faktor, který může ovlivnit abiotickou složku: ochlazování povrchů, odnos vlhkosti a zrychlení evaporace, transport a odnos menších částic různých materiálů (např v pouštích výrazným způsobem formuje utváření krajiny). Může ovlivnit i biotickou složku ekosystému: transport semen a pylu na větší vzdálenosti, obdobně ovlivnění pohybu hmyzu, nebo ptáků. Proudění větru zvyšuje i fyziologický výpar- transpiraci, čímž výrazně zasahuje do vodního režimu rostlin. V horských podmínkách částice unášené větrem mohou obrušovat návětrnou část dřevin a obrušovat na ní pupeny, díky čemuž vznikají tzv. vlajkové stromy.


Složení atmosféry v rámci biosféry se mění jen nepatrně: 21% kyslíku, 78% dusíku, 1% vzácné plyny a s nimi oxid uhličitý, jehož koncentrace může kolísat mezi létem a zimou (a převažující produkcí, nebo převažující spotřebou mezi 0,03-0,035%. Se vzrůstající nadmořskou výškou klesá hustota vzduchu (např. v 5000 m.n.m. je obsah kyslíku v m3 vzduchu poloviční, než na hladině moře), nemění se však poměr jednotlivých plynů.
Důležitou charakteristikou vzduchu, která se mění téměř neustále a těsně souvisí s průběhem počasí, je jeho vlhkost, udávající obsah vodních par(při dané teplotě a tlaku) a tlak vzduchu (přepočítaný na hladinu moře).
Vlhkost vzduchu
Absolutní vlhkost vzduchu
- je množství vody v g poutané v m3 vzduchu. Závisí na tlaku a teplotě vzduchu - čím je vzduch teplejší, tím větší množství vodních par v něm může být obsaženo.Relativní vlhkost vzduchu
-je poměr mezi skutečným a maximálně možným množstvím vodních par obsažených ve vzduchu za daných podmínek; udává se v %. Při překročení nasycení, na hodnotu větší než 100%, dochází ke kondenzaci (mlha, oblačnost). Kondenzace- srážení vodních par z plynného skupenství na kapalné - začíná probíhat na pevných površích (nejen větší plochy, ale i prachové částice v atmosféře) - nebo na předmětech, které jsou chladnější, než okolní vzduch. Relativní vlhkost vzduchu současně ukazuje i možnosti dalšího výparu- přechodu vody z kapalného skupenství na plynné - a z hlediska fungování rostlin i např. možnosti transpirace.Rosný bod
Vlhkost vzduchu tedy těsně souvisí s teplotou. Rosný bod je teplota, při které je vzduch maximálně nasycen vodními parami (relativní vlhkost vzduchu je 100 %). Pokud teplota klesne pod tento bod, nastává kondenzace. Přítomnost kondenzačních jader (pevná plocha, prachové částice) tuto kondenzaci urychluje, jejich nepřítomnost naopak kondenzaci oddálí i při podchlazení pod rosný bod. Teplota rosného bodu je různá pro různé absolutní vlhkosti vzduchu: čím větší je relativní vzdušná vlhkost, tím vyšší je teplota rosného bodu, a tím snadněji dojde při ochlazení ke kondenzaci.Tlak vzduchu
Je to tlak, kterým působí vzdušná vrstva na zemský povrch, závisí tedy mimo jiné i na nadmořské výšce (s výškou klesá). Dále závisí na teplotě, proudění větru a nasycení vzduchu vodními parami. Tlak se zjišťuje pomocí barometru, udává se v hPa (hektopascalech), přičemžnormální tlak = 1013 hPa. Na meteorologických mapách se tlak zachycuje ve formě Izobar - což jsou křivky, spojující místa se stejným tlakem. Tlakové níže jsou oblasti kde je tlak výrazně nižší než v okolí, u tlakových výší je tomu naopak.
Tlaková výše - Anticyklóna
Jedná se o oblast vyššího tlaku vzduchu v atmosféře, přičemž tlak vzduchu v jejím okolí je nižší než tlak uvnitř
oblasti. Chladný vzduch v ní svou vahou klesá dolů a dole se rozptyluje. Vlivem Coriolisovy síly se na severní polokouli
vzduch proudící z anticyklóny stáčí ve směru hodinových ručiček; na jižní polokouli proti směru hodinových ručiček.
Díky chladnému suchému vzduchu v místě Anticyklóny, který se ohřívá o zemský ovrch (a tím zvyšuje svou schopnost přijímat vodní páry, je v oblasti anticyklony velmi malá nebo žádná tvorba mraků.
Na synoptických mapách se anticyklóna značí písmenem V (nebo H) uzavřeným v izobaře.
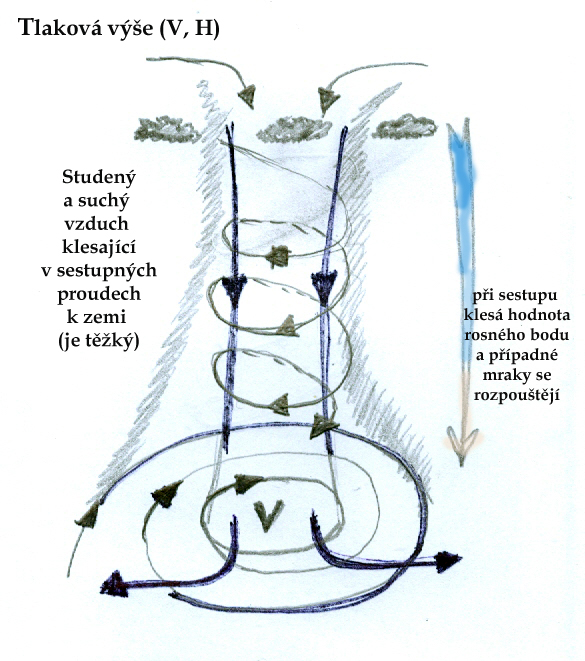
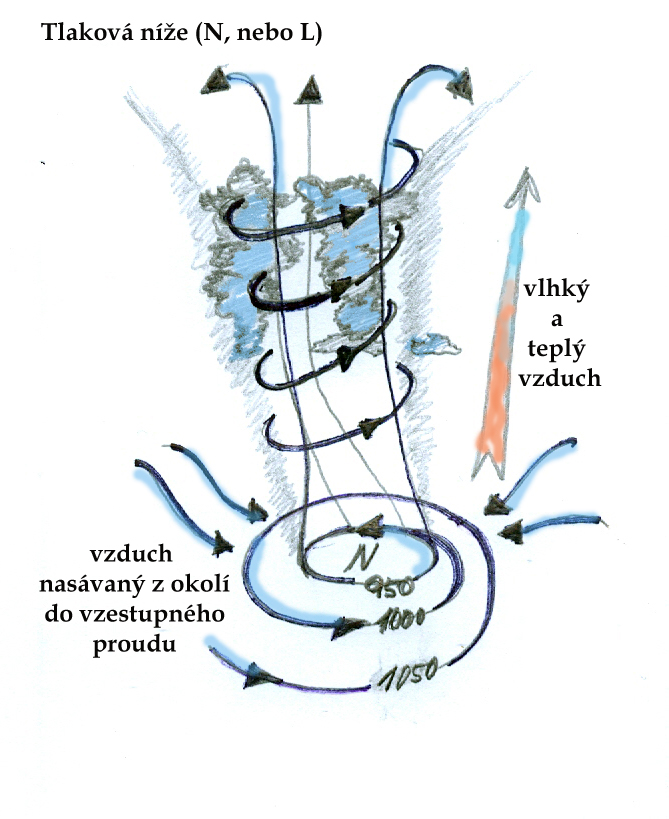
Tlaková níže - Cyklóna
Jedná se o oblast se sníženým tlakem vzduchu, přičemž tlak vzduchu v jejím okolí je vyšší než tlak uvnitř oblasti.
Teplý a vlhký vzduch v ní stoupá vzhůru díky komínovému efektu – svým výstupem se ochlazuje a vodní pára v něm obsažená
kondenzuje a vytváří mohutnou srážkově významnou oblačnost. Prvořadým zdrojem energie tohoto proudění (víru) je teplo uvolněné z kondenzujících vodních par.Vystupující vzduch se vlivem rotace Země na S polokouli stáčí vždy
proti směru pohybu hodinových ručiček,zatímco na J polokouli opačně.
Na synoptických mapách se značí písmenem N (nebo L) uzavřeným v izobaře
(někdy s udáním hodnoty tlaku).
Počasí
To, co vnímáme jako počasí, nebo změny počasí, je tedy soubor jevů v atmosféře, založený na fyzikálních zákonech a ovlivňovaný především následujícími faktory:
- Teploty vzdušných mas (vyšší hustotou a specifickou hmotností chladného vzduchu) a s tím souvisejícími vzestupnými nebo sestupnými proudy
- Snahou o vyrovnání teplot na navzájem ne příliš vzdálených místech vedoucí k proudění a jevu zvanému "vítr"
- Souvislost relativní vzdušné vlhkosti a jeho teploty, kdy při ochlazení vzduchu pod rosný bod dochází k tvorbě srážek
- Tlakem vzduchu působícího na zemských povrch- souvisejícího s ryhlostí a směrem proudění vzdušných mas
Některé z těchto jevů souvisí s kvalitou zemského povrchu, reliéfem, možnostmi výparu vody a obohacení vzduchu vlhkostí-
a proto je průběh počasí v některých oblastech stálejší- souvisí s ročními obdobími a například tím, jestli je oceán nebo pevnina v daném obdobéí teplejším povrchem
a jestli tedy bude převládající směr proudění z pevniny nad oceán, nebo naopak.
Změny počasí souvisí s přesuny různě chladných vzdušných mas a jejich vzájemným působením přinášejícím změnu počasí, oblačnosti a srážek. Místo styku různě teplých vzdušných mas, s výraznější změnou očasí, označujeme jako "fronty", nebo frontální poruchy.
Teplá fronta
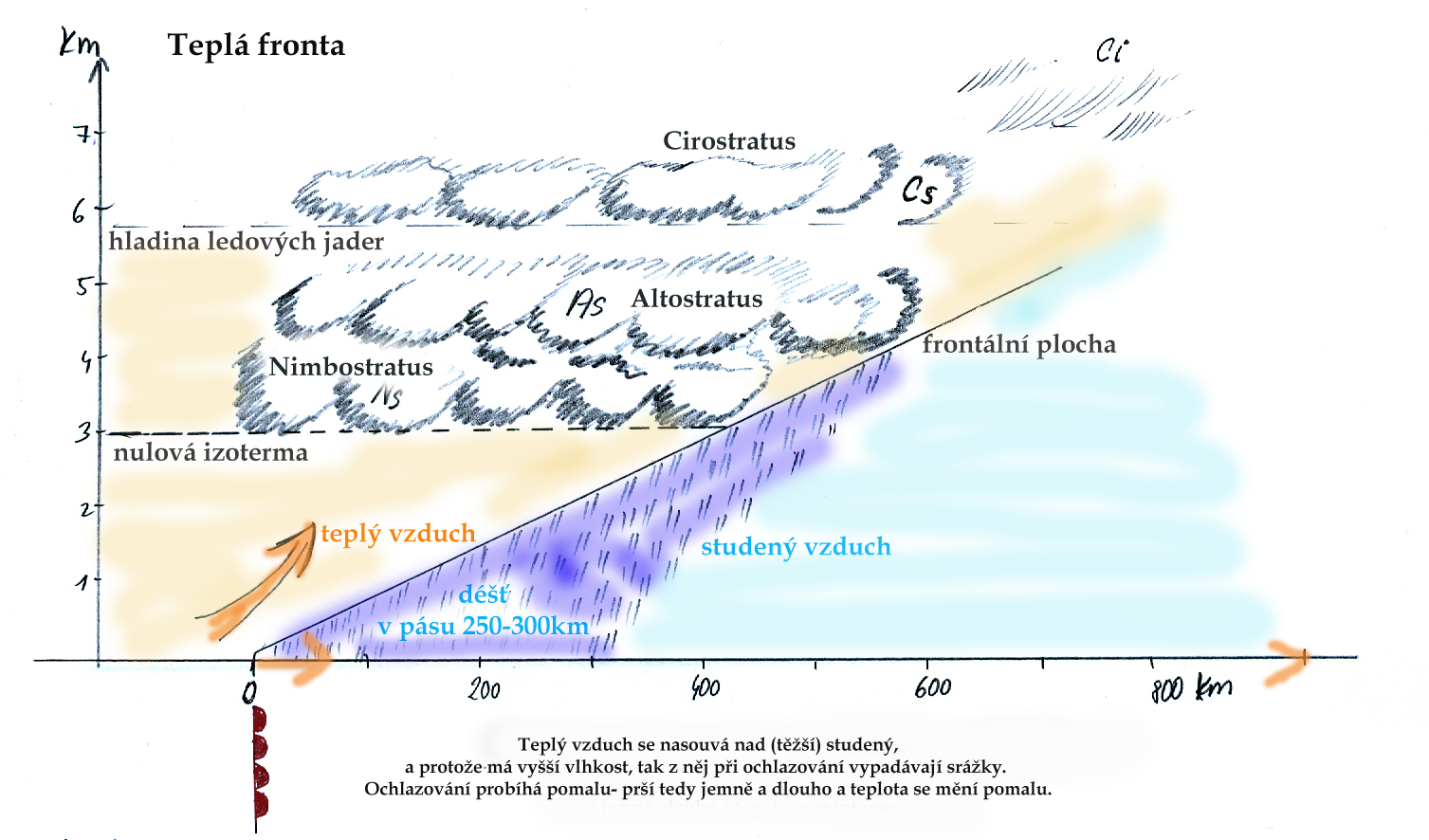
Teplá fronta je úzké rozhraní mezi studeným a teplým vzduchem, které se pohybuje směrem ke studenému vzduchu. Obvykle souvisí s výskytem tlakové níže.
Jestliže teplá (a vlhčí) vzduchová hmota proudí nad chladnější povrch, než byla oblast jejího vzniku,
v silných výstupních proudech dochází k jejímu postupnému ochlazení a postupné (ve výškovém gradientu) kondenzaci vzdušných par.
Na tomto rozhraní se pak vytváří mohutný systém typické vrstevnaté oblačnosti, která sahá stovky kilometrů před frontální čáru.
Vzniká zde mrholení a déšť trvalejšího charakteru a srážkové pásmo je obvykle několik stovek kilometrů široké.
Prvním příznakem blížící se teplé fronty jsou vysoké oblaky, snižující se viditelnost (popř. vzniká i mlha) a
klesající tlak vzduchu;
vítr se stáčí vlevo a zesiluje. Po přechodu teplé fronty trvalý déšť přestává, oblačnost se protrhává a zvedá.
Vítr se stáčí doprava a jeho síla klesá, tlak vzduchu se buď nemění, nebo nadále zvolna klesá.
Za teplou frontou převládá teplé počasí, otepluje se.
Pohyb teplé fronty je relativně pomalý.
Studená fronta
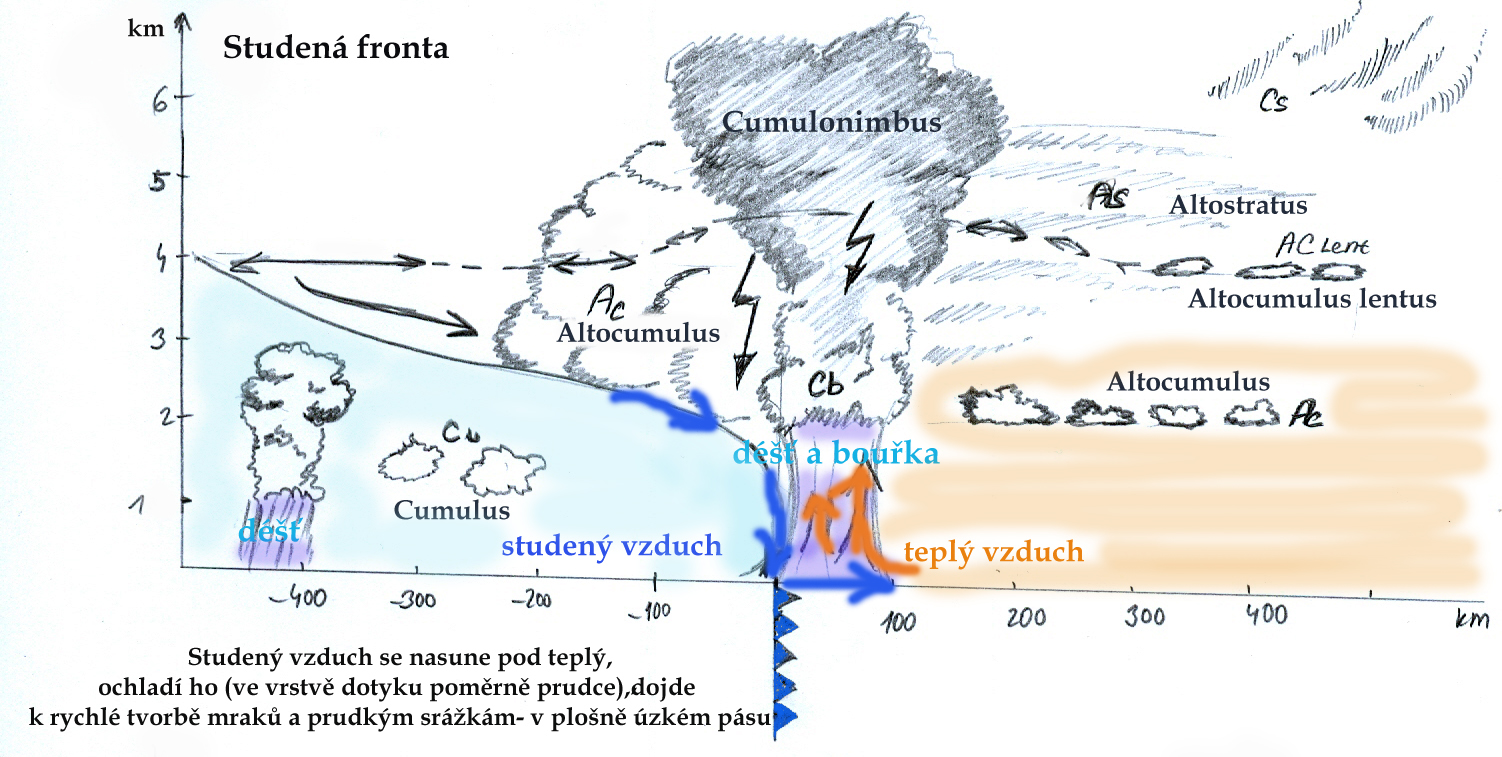
je úzké rozhraní mezi teplým a studeným vzduchem, které se pohybuje směrem k teplému vzduchu.
Obvykle uzavírá teplý sektor tlakové níže.
Jestliže chladnější (a sušší) vzduchová hmota proudí nad teplejší povrch, než byla oblast jejího vzniku,
nasouvá se (díky vyšší hustotě chladného vzduchu) pod teplý vduch v daném místě.
a dojde v ní k výstupným pohybům vzduchu (jsou mohutnější než v teplé frontě), vynášejícím do velkých výšek vlhký teplý vzduch.
Vlhkost zde vypadává ve formě srážek, ale íky silným vzestupným proudům může v mracích cirkulovat, kapky fungují jako kondenzační jádra a nabalují na sebe další vlhkost-
a pokud k jevu dochází nad hranicí ledových jader, mohou vznikat i kroupy.
Studená fronta se většinou projevuje kupovitou oblačností s přeháňkami, provázenými v létě i bouřkami.
Srážkové pásmo má užší, než teplá fronta (ca 100-150km)a srážky se vyskytují i před frontální čarou.
Typickými jevy jsou oblaka druhu Cumulonimbus, silné deště (lijáky), intenzivní bouřky a prudké nárazy větru převyšující někdy 10 stupňů Beauforta.
Vítr se před frontou stáčí doleva, na frontě se uklidní a po přechodu fronty se rychle stáčí doprava.
Tlak vzduchu před studenou frontou klesá, za ní výrazně stoupá.
Rychlost pohybu studené fronty je obvykle vyšší, než rychlost pohybu fronty teplé (ca o 20-50%).)
Okluzní fronta
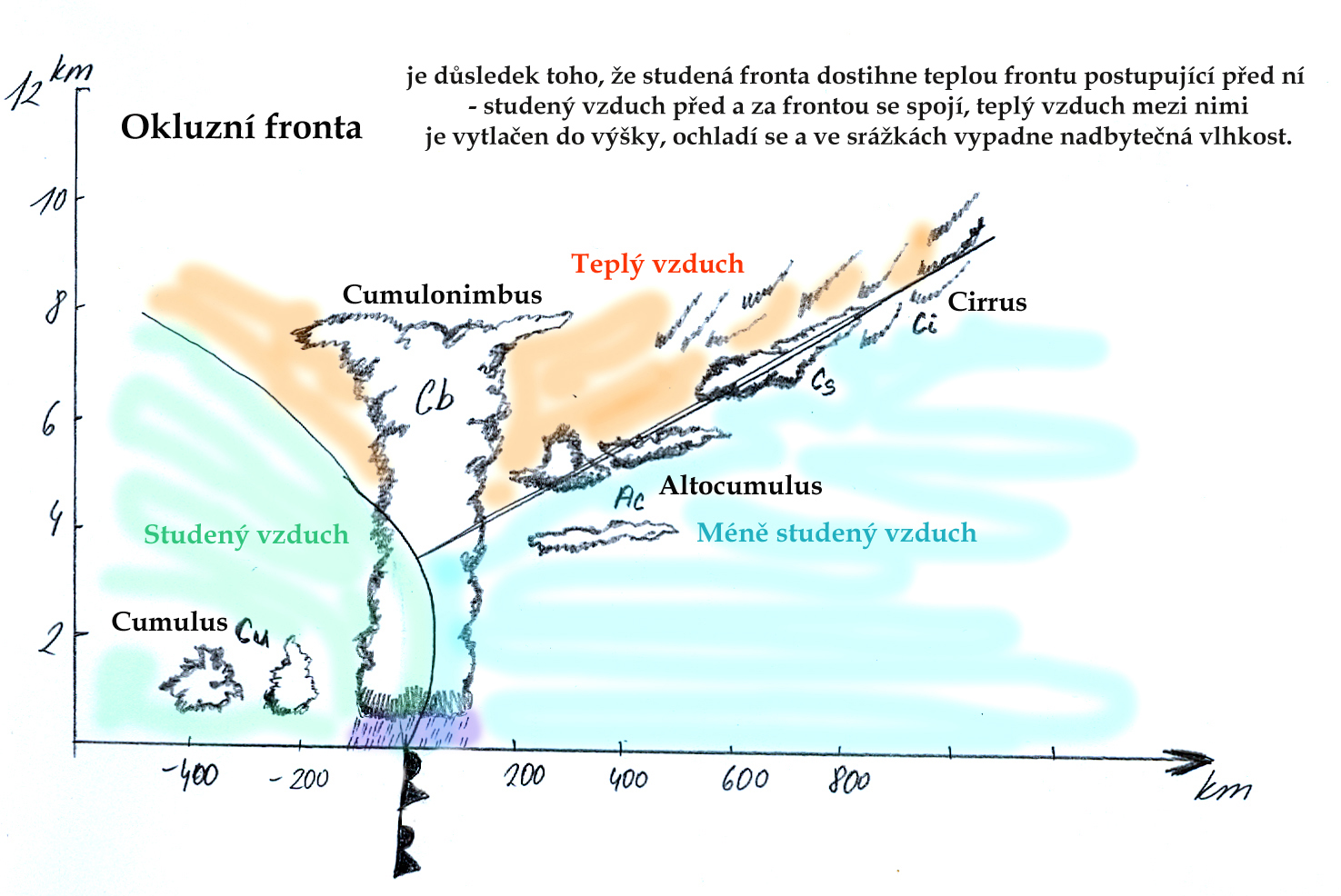
Okluzní fronta vzniká za situace, kdy studená fronta dostihne teplou frontu postupující před ní a u zemského povrchu se spojí studený vzduch za studenou frontou, s (o něco méně) studeným vzduchem před teplou frontou a teplý vzduch je oběma chladnými vzdušnými masami současně vytlačen do výše. Oblast styku dřívější teplé a studené fronty dostala název okluzní fronta, podle procesu uzavírání teplého sektoru tlakové níže následnou frontou (okluze). Počasí na okluzních frontách je nevýrazné, hůře předvídatelné a podobá se buďto počasí typickému pro studenou frontu, nebo pro teplou frontu; intenzita větru i srážky bývají však většinou slabší, než na obou těchto frontách.
V našich podmínkách tlaková níže často přináší větrné a deštivé počasí.
Tlaková výše v létě přináší minimální oblačnost, žádné srážky a nanejvýš slabý vítr (anticyklonální počasí). V zimním období může přinést buďto jasné a mrazivé počasí beze srážek, nebo naopak vést k tvorbě "inverzí": pod tzv. inverzní vrstvou je chladný vzduch, obsahující kondenzační jádra a vzniká v něm mlha a srážky, nad inverzní vrstvou je jasno a vzduch je ohříván sluncem, tudíž v něm nedochází ke kondenzaci. Obě vrstvy se navzájem nepromíchávají.
Kromě krátkodobých anticyklón se na Zemi vyskytuje řada anticyklón sezónního či permanentního charakteru. Ze stálých anticyklón má vliv na počasí v Evropě především anticyklóna Azorská, na jižní polokouli anticyklóna Antarktická. Ze sezónních anticyklón vynikají kontinentální anticyklóny: Sibiřská vytvářející se v zimním období nad promrzlými částmi Asie (střed nad Mongolskem) a anticyklóna Kanadská (se středem nad Skalistými horami a Velkými kanadskými jezery)
Literatura použitá a doporučená:
- Baier J., Baierová V., 1985: Abeceda výživy rostlin a hnojení. 1. vyd. Praha, Státní zemědělské nakladatelství. 364 s.
- Begon M., Harper l. J., Townsend C.R.: Ekologie- jedinci, populace a společenstva, Vydavatelství Univerzity Palackého Olomouc, 1997, s.950
- Hartmann G., Nienhaus F., Butin H.,2001 : Atlas poškození lesních dřevin, Brázda s.r.o. Praha, , 287 s.
- Chroust L.1997: Ekologie výchovy lesních porostů, Opočno, FGMRI RS Opočno. 277 s.
- Jandák J., Prax A., Pokorný E.,12001: Půdoznalství, MENDELU Brno. 140s.
- Jeník J.1995: Ekosystémy. /Ecosystems./ Praha, UK . 135 s.
- Klimo, E.1994: Ekologie lesa. /Ekology of the Forest/. Brno, VŠZ v Brně. 170 s.
- Korpel´ Š.1991: Pestovanie lesa. /Silviculture/. Príroda, Bratislava. 465 s.
- Korpel Š. , Saniga, M.1995: Prírode blízke pestovanie lesa. Zvolen, ÚVVP LVH SR Zvolen. 158 s.
- Kulhavý a kol., 2003: Ekologie lesa I, Učební text MZLU v Brně: 220 s.
- Kulhavý a kol.2006: Ekologie lesa II, Učební text MZLU v Brně: 221 s.
- Kulhavý a kol. 2009: Ekologie lesa III, Učební text MZLU v Brně: 120 s.
- Míchal I.,1994: Ekologická stabilita. Veronica, Brno. 275s.
- Míchal I. et al.,1992: Obnova ekologické stability lesů. Praha, Academia. 169 s.
- Poleno Z., Vacek S.,2007: Pěstování lesů I. - Ekologické základy pěstování lesů, Lesnická práce: 316 s.
- Prax A., Pokorný E.,1996: Klasifikace a ochrana půd, MENDELU Brno. 143s.
- Procházka S. a kol, 1998: Fyziologie rostlin, Academia Praha, 485s.
- Rosypal S. a kol, 1994: Přehled biologie, Scientia s.r.o., 1994, s.578.
- Šarman, J., 1981: Lesnické půdoznalství s mikrobiologií. Praha, Státní pedagogické nakladatelství. 225 s.
- Volf F. a kol.,1988: Zemědělská botanika, SZN. 383 s.
- Vodrážka Z.,1999: Biochemie, Academia Praha. 191s.
- http://klasifikace.pedologie.czu.cz/index.php?action=showKlasifikacniSystem
- ftp://ftp.uhul.cz/public/typologie/taxonomicky_klasifikacni_system_pud_v_cr.pdf